
Journal of Tropical Oceanography >
A study of shell-selection preference of two species hermit crab in the Xisha Islands
Received date: 2023-11-26
Revised date: 2024-01-31
Online published: 2024-03-05
Supported by
Open Project of Yazhou Bay Innovation Research Institute, Hainan Tropical Ocean University(2022CXYKFKT07)
National Natural Science Foundation of China(32002389)
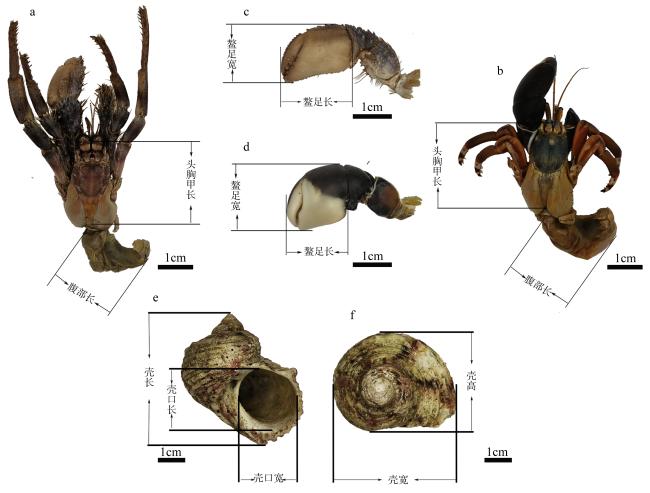
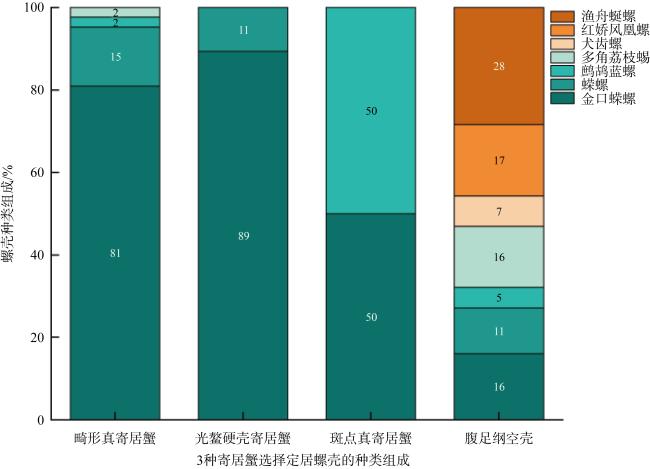
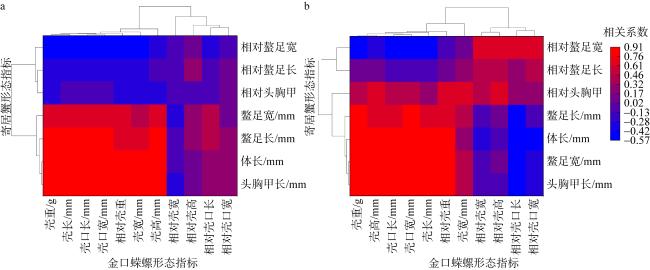
To investigate the behavior and related factors of hermit crabs in selecting shells for settlement, this study collected 81 empty gastropod shells and 72 hermit crab samples from the intertidal zone of the Xisha Islands. Results obtained through morphometric measurements and statistical methods on two species of hermit crabs and the shells of Turbo argyrostomus show that in the environments where the types and abundance of shells are relatively rich, both Dardanus deformi and Calcinus laevimanus preferentially choose shells from the family Turbinidae for settlement. Specifically, the shells of T. argyrostomus are more likely to become settlement sites for both hermit crab species. Statistical analysis of morphological parameters reveals significant differences between the two species in shell weight, shell length, shell width, shell height, aperture length, and aperture width during shell selection. Standardized data indicate differences in shell size preferences between the two hermit crab species. D. deformi primarily considers the ratio of cephalothoracic length to shell length, while C. laevimanus focuses on the ratios of cheliped width to shell width, shell height, and aperture length. In summary, this study provides an in-depth exploration of the complex process of hermit crab shell selection, revealing differences in shell preferences among different hermit crab species. The findings contribute valuable insights into the ecological behavior of hermit crabs, aiding in understanding their roles and adaptive mechanisms in intertidal ecosystems.
WANG Haishan , CHEN Zhi , LI Youming , YE Le . A study of shell-selection preference of two species hermit crab in the Xisha Islands[J]. Journal of Tropical Oceanography, 2024 , 43(5) : 84 -92 . DOI: 10.11978/2023176
表1 两种寄居蟹及其定居螺壳形态描述性统计Tab. 1 Descriptive statistics on the morphology of two species of hermit crabs and their sedentary conch shells |
| 物种 | 形态参数 | 畸形真寄居蟹 | 光螯硬壳寄居蟹 | ||||
|---|---|---|---|---|---|---|---|
| 均值 | 标准差 | 变异系数 | 均值 | 标准差 | 变异系数 | ||
| 金口 蝾螺 | 壳重/g | 28.58 | 10.51 | 0.37 | 18.33 | 3.84 | 0.21 |
| 壳长/mm | 47.77 | 6.85 | 0.14 | 40.29 | 6.63 | 0.16 | |
| 壳宽/mm | 42.85 | 8.63 | 0.20 | 39.43 | 5.12 | 0.13 | |
| 壳高/mm | 31.37 | 5.66 | 0.18 | 25.43 | 2.48 | 0.10 | |
| 壳口长/mm | 26.92 | 4.67 | 0.17 | 23.88 | 2.26 | 0.09 | |
| 壳口宽/mm | 22.48 | 3.98 | 0.18 | 19.51 | 2.26 | 0.12 | |
| 相对壳重 | 0.58 | 0.15 | 0.26 | 0.46 | 0.07 | 0.15 | |
| 相对壳宽 | 0.90 | 0.14 | 0.16 | 0.99 | 0.14 | 0.14 | |
| 相对壳高 | 0.66 | 0.08 | 0.12 | 0.64 | 0.06 | 0.09 | |
| 相对壳口长 | 0.56 | 0.04 | 0.07 | 0.60 | 0.08 | 0.14 | |
| 相对壳口宽 | 0.47 | 0.03 | 0.07 | 0.49 | 0.06 | 0.12 | |
| 寄居蟹 | 头胸甲长/mm | 25.18 | 4.66 | 0.19 | 19.51 | 2.85 | 0.15 |
| 体长/mm | 48.90 | 9.99 | 0.20 | 37.59 | 6.66 | 0.18 | |
| 螯足长/mm | 20.32 | 3.90 | 0.19 | 15.79 | 3.47 | 0.22 | |
| 螯足宽/mm | 13.40 | 1.94 | 0.14 | 11.06 | 2.25 | 0.20 | |
| 相对头胸甲长 | 0.52 | 0.04 | 0.07 | 0.52 | 0.04 | 0.07 | |
| 相对螯足长 | 0.42 | 0.04 | 0.11 | 0.42 | 0.09 | 0.20 | |
| 相对螯足宽 | 0.28 | 0.04 | 0.13 | 0.30 | 0.06 | 0.21 | |
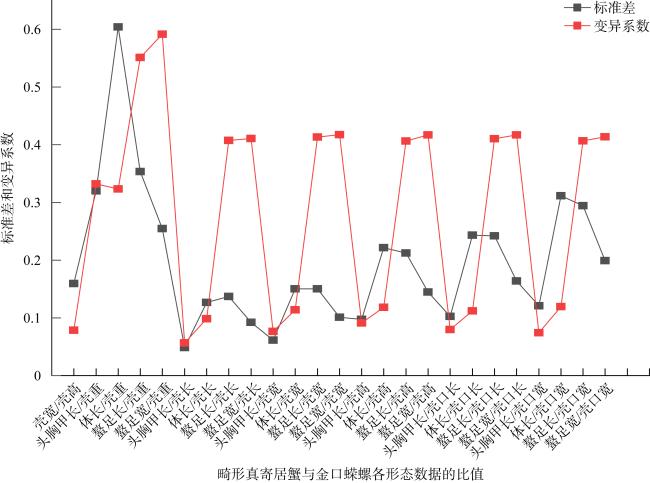
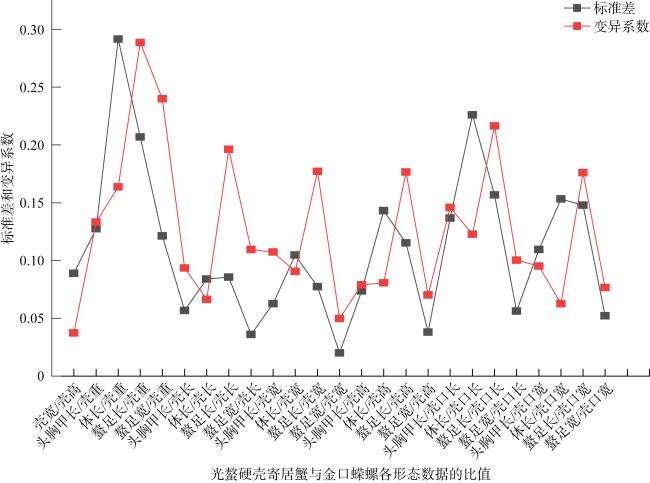
表2 寄居蟹与螺壳形态指标比值统计Tab. 2 Statistics on the ratio of morphological indexes between hermit crabs and snail shells |
| 形态比值 | 畸形真寄居蟹 | 光螯硬壳寄居蟹 | ||||
|---|---|---|---|---|---|---|
| 均值 | 标准差 | 变异系数 | 均值 | 标准差 | 变异系数 | |
| 壳宽/壳高 | 1.4450 | 0.1596 | 0.1105 | 1.4959 | 0.0890 | 0.0595 |
| 头胸甲长/壳重 | 1.0473 | 0.3200 | 0.3056 | 0.8621 | 0.1275 | 0.1479 |
| 体长/壳重 | 2.0222 | 0.6041 | 0.2987 | 1.6564 | 0.2916 | 0.1761 |
| 螯足长/壳重 | 0.7459 | 0.3536 | 0.4741 | 0.7096 | 0.2069 | 0.2915 |
| 螯足宽/壳重 | 0.5039 | 0.2545 | 0.5050 | 0.4925 | 0.1213 | 0.2464 |
| 头胸甲长/壳长 | 0.5192 | 0.0485 | 0.0935 | 0.5090 | 0.0567 | 0.1114 |
| 体长/壳长 | 1.0076 | 0.1268 | 0.1258 | 0.9718 | 0.0839 | 0.0863 |
| 螯足长/壳长 | 0.3776 | 0.1372 | 0.3635 | 0.4153 | 0.0856 | 0.2061 |
| 螯足宽/壳长 | 0.2518 | 0.0922 | 0.3659 | 0.2867 | 0.0361 | 0.1260 |
| 头胸甲长/壳宽 | 0.5628 | 0.0612 | 0.1087 | 0.5054 | 0.0627 | 0.1241 |
| 体长/壳宽 | 1.0921 | 0.1503 | 0.1376 | 0.9657 | 0.1048 | 0.1086 |
| 螯足长/壳宽 | 0.4080 | 0.1501 | 0.3679 | 0.4106 | 0.0774 | 0.1885 |
| 螯足宽/壳宽 | 0.2722 | 0.1010 | 0.3710 | 0.2829 | 0.0201 | 0.0709 |
| 头胸甲长/壳高 | 0.8096 | 0.0973 | 0.1202 | 0.7530 | 0.0737 | 0.0979 |
| 体长/壳高 | 1.5695 | 0.2214 | 0.1410 | 1.4411 | 0.1432 | 0.0994 |
| 螯足长/壳高 | 0.5851 | 0.2123 | 0.3627 | 0.6132 | 0.1152 | 0.1879 |
| 螯足宽/壳高 | 0.3909 | 0.1449 | 0.3707 | 0.4232 | 0.0380 | 0.0898 |
| 头胸甲长/壳口长 | 0.9206 | 0.1024 | 0.1112 | 0.8559 | 0.1367 | 0.1597 |
| 体长/壳口长 | 1.7859 | 0.2432 | 0.1362 | 1.6339 | 0.2260 | 0.1383 |
| 螯足长/壳口长 | 0.6618 | 0.2420 | 0.3658 | 0.6974 | 0.1567 | 0.2247 |
| 螯足宽/壳口长 | 0.4417 | 0.1638 | 0.3709 | 0.4788 | 0.0563 | 0.1176 |
| 头胸甲长/壳口宽 | 1.1299 | 0.1209 | 0.1070 | 0.9699 | 0.1095 | 0.1129 |
| 体长/壳口宽 | 2.1943 | 0.3115 | 0.1419 | 1.8514 | 0.1533 | 0.0828 |
| 螯足长/壳口宽 | 0.8106 | 0.2943 | 0.3630 | 0.7890 | 0.1479 | 0.1875 |
| 螯足宽/壳口宽 | 0.5412 | 0.1993 | 0.3682 | 0.5447 | 0.0521 | 0.0957 |
| [1] |
贾男, 周天成, 胡思敏, 等, 2024. 南沙群岛海域珊瑚礁区三种寄居蟹的摄食差异比较[J]. 热带海洋学报, 43(3): 109-121.
|
| [2] |
荆圆圆, 胡凡光, 刘广斌, 等, 2023. 山东沿海不同地理群体中国蛤蜊形态差异分析[J]. 海洋渔业, 45(1): 95-104.
|
| [3] |
王旭, 徐衡, 邹莉, 等, 2013. 3个单齿螺地理群体形态差异分析[J]. 南方水产科学, 9(4): 22-27.
|
| [4] |
张素萍, 2008. 中国海洋贝类图鉴[M]. 北京: 海洋出版社: 40.
|
| [5] |
周天成, 2020. 三亚鹿回头近岸珊瑚礁区寄居蟹的摄食及生态功能探讨[D]. 北京: 中国科学院大学: 1-18.
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}