
Journal of Tropical Oceanography >
Taxonomic study of four species of Gracilaria (Gracilariaceae, Rhodophyta) in Zhanjiang based on morphological and molecular data
Copy editor: YIN Bo
Received date: 2023-06-26
Revised date: 2023-09-03
Online published: 2024-03-26
Supported by
Project of Southern Marine Science and Engineering Guangdong Laboratory (Zhanjiang)(ZJW-2022-06)
Special Science and Technology Innovation Project of Guangdong Province, China(2022A01207)
Ph.D. Start-up Fund of Guangdong Medical University, China(GDMUB2022025)
Discipline Construction Project of Guangdong Medical University, China(4SG23009G)
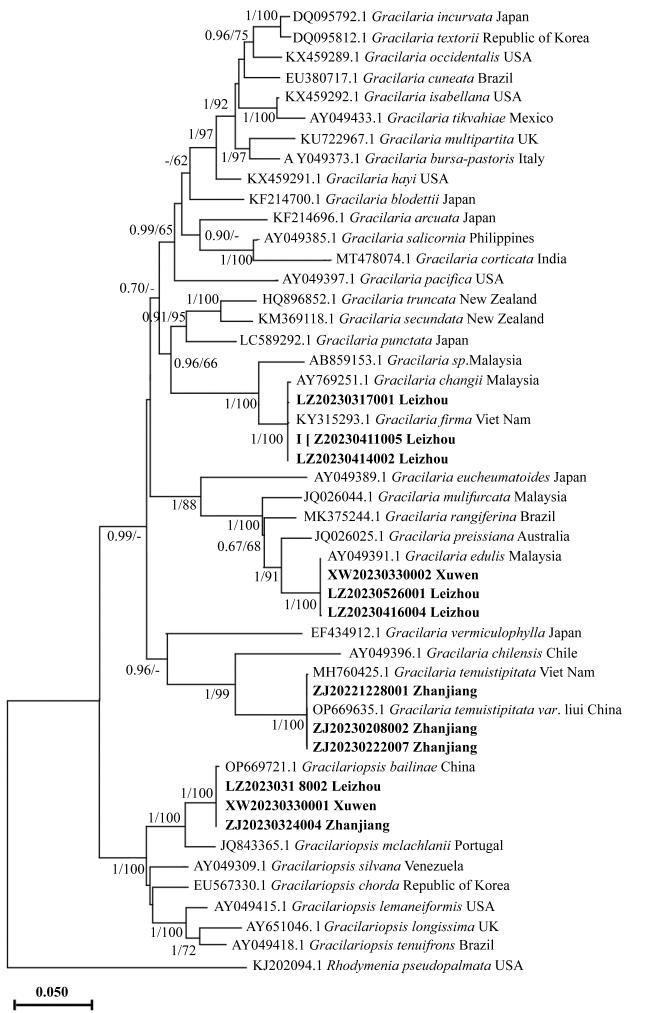
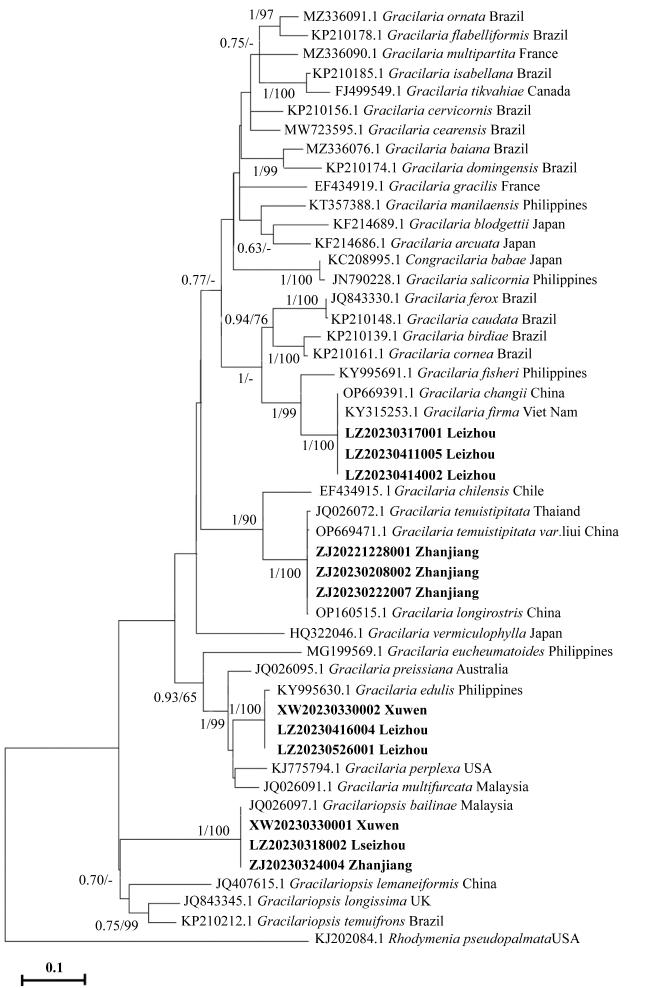
According to the molecular and morphological analysis, the species of Gracilariaceae was studied using the samples collected from Zhanjiang, Guangdong province, China. Four Gracilariaceae species were identified, namely: Gracilaria tenuistipitata, G. firma, G. edulis and Gracilariopsis bailinae. Their taxonomic characters and phylogenetic relationships were discussed in detail. This is combined with data from the literature to clarify that G. tenuistipitata, G. tenuistipitata var. liui and G. longirostris are the same species, and G. firma and G. changii are the same species. This study revised the taxonomic information of Gracilariaceae species in the coastal waters of southern China, and also provided a taxonomic basis for the conservation and development of macroalgae resources in Zhanjiang sea area.
Key words: Zhanjiang sea area; morphology; Gracilaria; phylogenetic analysis
LI Nenghui , HUANG Qing , LI Hang , ZENG Jun , WU Kefeng , TAN Huaqiang . Taxonomic study of four species of Gracilaria (Gracilariaceae, Rhodophyta) in Zhanjiang based on morphological and molecular data[J]. Journal of Tropical Oceanography, 2024 , 43(2) : 34 -47 . DOI: 10.11978/2023085
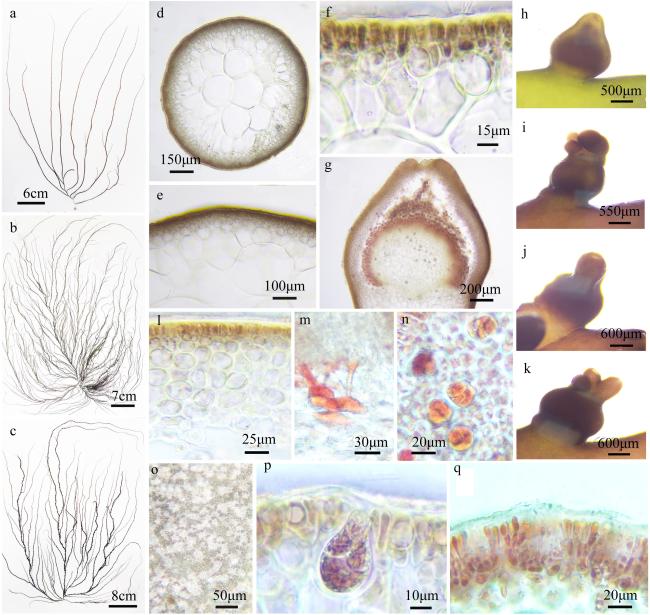
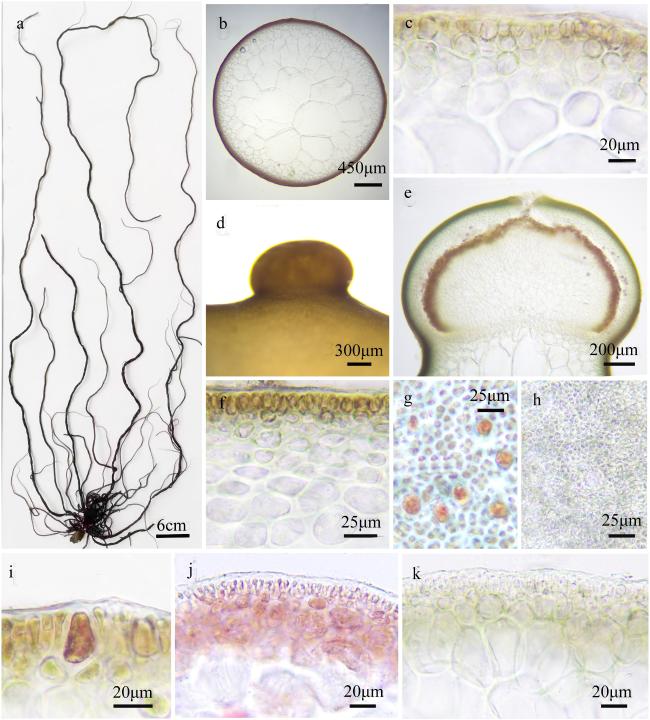
图2 细基江蓠a~c. 藻体外观; d~f. 藻体横切面观; g. 囊果切面观; h~k. 囊果外观; l. 囊果被切面观; m. 吸收丝; n. 四分孢子囊表面观; o. 精子囊窠表面观; p. 四分孢子囊切面观; q. 精子囊窠切面观 Fig. 2 Gracilaria tenuistipitata. a ~ c. The habit sketch of the frond; d ~ f. The transection of the frond; g. the longitudinal section of the cystocarp; h ~ k. The surface of the cystocarp; l. The longitudinal section of the pericarp; m. the traversing filaments; n. The surface of the tetrasporangia; o. The surface of the spermatangia; p. The transection of the tetrasporangia; q. The transection of the spermatangia |
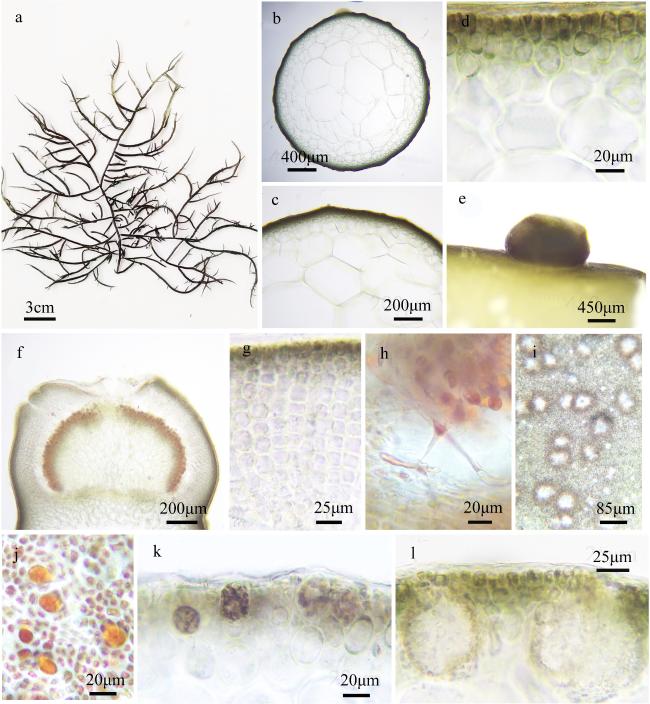
图3 硬江蓠a. 藻体外形图; b~d. 藻体横切面观; e. 囊果外观; f. 囊果切面观; g. 囊果被切面观; h. 吸收丝; i. 精子囊窠表面观; j. 四分孢子囊表面观; k. 四分孢子囊切面观; l. 精子囊窠切面观 Fig. 3 Gracilaria firma. a. The habit sketch of the frond; b ~ d. The transection of the frond; e. The surface of the cystocarp; f. The longitudinal section of the cystocarp; g. The longitudinal section of the pericarp; h. The traversing filaments; i. The surface of the spermatangia; j. The surface of the tetrasporangia; k. The transection of the tetrasporangia; l. The transection of the spermatangia |
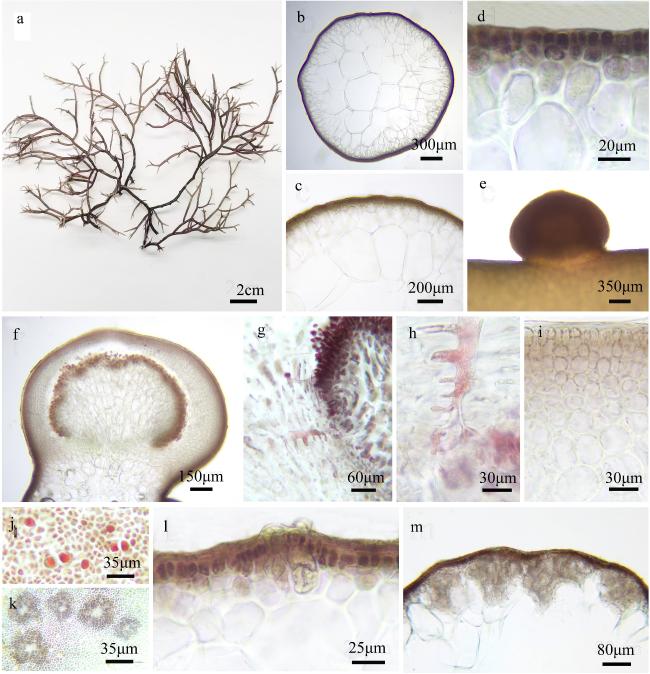
图4 帚状江蓠a. 藻体外形图; b~d. 藻体横切面观; e. 囊果外观; f. 囊果切面观; g~h. 吸收丝; i. 囊果被切面观; j. 四分孢子囊表面观; k. 精子囊窠表面观; l. 四分孢子囊切面观; m. 精子囊窠切面观 Fig. 4 Gracilaria edulis. a. The habit sketch of the frond; b ~ d. The transection of the frond; e. The surface of the cystocarp; f. The longitudinal section of the cystocarp; g ~ h. The traversing filaments; i. The longitudinal section of the pericarp; j. The surface of the tetrasporangia; k. The surface of the spermatangia; l. The transection of the tetrasporangia; m. The transection of the spermatangia |
图5 异枝拟江蓠a. 藻体外形图; b~c. 藻体横切面观; d. 囊果外观; e. 囊果切面观; f. 囊果被切面观; g. 四分孢子囊表面观; h. 精子囊表面观; i. 四分孢子囊切面观; j~k. 精子囊切面观 Fig. 5 Gracilariopsis bailinae. a. The habit sketch of the frond; b ~ c. The transection of the frond; d. The surface of the cystocarp; e. The longitudinal section of the cystocarp; f. The longitudinal section of the pericarp; g. The surface of the tetrasporangia; h. The surface of the spermatangia; i. The transection of the tetrasporangia; j ~ k. The transection of the spermatangia |
表1 样品与细基江蓠及其繁枝变种的形态特征比较Tab. 1 Comparison of morphological characteristics of the samples with G. tenuistipitata and G. tenuistipitata var. liui |
| 结构 | 样品(分枝简单) | 样品(分枝繁多) | 细基江蓠 | 细基江蓠繁枝变种 |
|---|---|---|---|---|
| 高 | 20~100cm | 20~85cm | 20~40cm | 20~30cm |
| 宽 | 0.5~3.6mm | 0.5~3mm | 0.5~1mm | 0.3~0.5mm |
| 表层细胞 | 1~2层; 5.8~19.6μm × 4.2~11μm | 1~2层; 5.8~17.5μm × 5~13μm | 1~2层; 15~18μm × 10.5~19.5μm | 1~2层; 12~18μm × 10~11μm |
| 内皮层细胞 | 2~3层; 23.1~84.6μm | 2~3层; 23.3~80.4μm | 2~3层; 33~83μm | 2~4层; 25~80μm |
| 髓部细胞 | 185~501μm | 190~485μm | 225~390μm | 200~250μm |
| 囊果 | 595~1780μm | 580~1830μm | 830~950μm | 0.8~1mm |
| 囊果被 | 6~13层; 厚72~185μm | 6~11层; 厚74~200μm | 8~11层; 厚72~102μm | 7~10层; 厚72~102μm |
| 吸收丝 | 存在 | 存在 | 存在 | 存在 |
| 精子囊 | “T”型, 24~50μm × 20~42μm | “T”型, 26~52μm × 18~40μm | “T”型, 19~23μm × 10~23μm | “T”型, 19~23μm × 10~23μm |
| 四分孢子囊 | 26.8~50.5μm × 18.4~43.2μm | 28.5~51.2μm × 16.7~42.5μm | 30~46μm × 18~30μm | 30~46μm × 18~30μm |
| 参考文献 | 本文 | 本文 | 张峻甫等(1976); 夏邦美(1999) | 夏邦美(1999); 丁兰平等(2014) |
表2 样品与细基江蓠及长喙江蓠的形态特征比较Tab. 2 Comparison of morphological characteristics of the samples with G. tenuistipitata and G. longirostris |
| 结构 | 样品(分枝简单) | 样品(囊果长喙) | 细基江蓠 | 长喙江蓠 |
|---|---|---|---|---|
| 高 | 20~100cm | 20~74cm | 20~40cm | 约20cm |
| 宽 | 0.5~3.6mm | 0.5~2.8mm | 0.5~1mm | 约 1mm |
| 表层细胞 | 1~2层; 5.8~19.6μm × 4.2~11μm | 1~2层; 6~18.5μm × 4.5~12.5μm | 1~2层; 15~18μm × 10.5~19.5μm | 1~2层; 8~13μm × 7~9μm |
| 内皮层细胞 | 2~3层; 23.1~84.6μm | 2~3层; 20~78.5μm | 2~3层; 33~83μm | 1~2层; 15~40μm |
| 髓部细胞 | 185~501μm | 185~470μm | 225~390μm | 70~200μm |
| 囊果 | 595~1780μm | 喙长1.1mm左右 | 830~950μm | 喙长2~3mm |
| 囊果被 | 6~13层; 厚72~185μm | 6~10层; 厚78~180μm | 8~11层; 厚72~102μm | 4~7层; 厚106~145μm |
| 吸收丝 | 存在 | 存在 | 存在 | 存在 |
| 精子囊 | “T”型, 24~50μm × 20~42μm | 未发现 | “T”型, 19~23μm × 10~23μm | 未发现 |
| 四分孢子囊 | 26.8~50.5μm × 18.4~43.2μm | 未发现 | 30~46μm × 18~30μm | 20~50μm × 10~36μm |
| 参考文献 | 本文 | 本文 | 张峻甫等(1976); 夏邦美(1999) | 夏邦美(1999); 丁兰平等(2014) |
表3 样品与张氏江蓠及硬江蓠的形态特征比较Tab. 3 Comparison of morphological characteristics of the samples with G. changii and G. firma |
| 结构 | 样品 | 张氏江蓠 | 硬江蓠 |
|---|---|---|---|
| 高 | 10~28cm | 7~14cm | 10~20cm |
| 宽 | 1.5~4.2mm | 0.5~2mm | 1~3mm |
| 表层细胞 | 1~2层; 6.6~13.2μm × 5.2~8.3μm | 1~2层; 6.6~9.9μm × 4~6.6μm | 2~3层; 6.6~13.2μm × 3.3~6.6μm |
| 内皮层细胞 | 2~4层; 13~45μm | 2~3层; 33~83μm | 3~5层; 26~83μm |
| 髓部细胞 | 300~800μm | 300~499μm | 230~450μm |
| 囊果 | 573~1161μm × 778~1600μm | 589~664μm × 930~996μm | 580~630μm |
| 囊果被 | 9~13层; 厚108~180μm | 10~12层; 厚79~132μm | 8~10层; 厚83~95μm |
| 吸收丝 | 存在 | 存在 | 不存在 |
| 精子囊 | “V”型, 56~102μm × 40~90μm | “V”型, 50μm × 33~36μm; “P”型, 69~76μm × 43~50μm | “V”型, 66~116μm × 33~66μm |
| 四分孢子囊 | 23~47μm × 13~29μm | 10~66μm × 20~40μm | 50~66μm × 26~43μm |
| 参考文献 | 本文 | Zhang等(1991) | 张峻甫等(1976) |
| [1] |
丁兰平, 黄冰心, 谢艳齐, 2011. 中国大型海藻的研究现状及其存在的问题[J]. 生物多样性, 19(6): 798-804.
|
| [2] |
丁兰平, 谭华强, 孙国栋, 等, 2014. 粤东地区江蓠属Gracilaria (江蓠科, 红藻门)的形态分类学[J]. 热带海洋学报, 33(4): 33-39.
|
| [3] |
丁兰平, 黄冰心, 王宏伟, 2015. 中国海洋红藻门新分类系统[J]. 广西科学, 22(2): 164-188.
|
| [4] |
丁兰平, 杨楠, 刘金梅, 等, 2020. 海洋经济红藻江蓠科的分类研究现状及存在的问题[J]. 应用海洋学学报, 39(1): 144-147.
|
| [5] |
李方洲, 黄亚冬, 郭永军, 等, 2022. 不同饵料对皱纹盘鲍养殖效果的影响[J]. 饲料研究, 45(7): 56-61.
|
| [6] |
李来好, 李刘冬, 杨贤庆, 等, 2003. 江蓠方便食品的加工工艺及毒理的研究[J]. 食品与发酵工业, 29(7): 10-13.
|
| [7] |
李文红, 胡自民, 覃志彪, 等, 2004. 细基江蓠及其繁枝变种的RAPD和ITS分析[J]. 海洋学报, 26(6): 89-95.
|
| [8] |
刘思俭, 2001. 我国江蓠的种类和人工栽培[J]. 湛江海洋大学学报, 21(3): 71-79 (in Chinese).
|
| [9] |
潘江球, 李思东, 2010. 江蓠的资源开发利用新进展[J]. 热带农业科学, 30(10): 47-50.
|
| [10] |
夏邦美, 1999. 中国海藻志第二卷红藻门第五册伊谷藻目杉藻目红皮藻目[M]. 北京: 科学出版社 (in Chinese).
|
| [11] |
谢恩义, 申玉春, 叶宁, 等, 2009. 流沙湾的底栖大型海藻调查[J]. 广东海洋大学学报, 29(4): 30-35.
|
| [12] |
杨宇峰, 费修绠, 2003. 大型海藻对富营养化海水养殖区生物修复的研究与展望[J]. 青岛海洋大学学报, 33(1): 53-57.
|
| [13] |
张才学, 周伟男, 孙省利, 等, 2020. 硇洲岛大型海藻群落的季节演替[J]. 热带海洋学报, 39(1): 74-84.
|
| [14] |
张峻甫, 夏邦美, 1962. 中国江蓠属植物地理学的初步研究[J]. 海洋与湖沼, 4(2): 189-198.
|
| [15] |
张峻甫, 夏邦美, 1963. 江蓠科的一个新属—多穴藻属[J]. 海洋科学集刊, 3: 119-126 (in Chinese).
|
| [16] |
张峻甫, 夏邦美, 1976. 中国江蓠属海藻的分类研究[J]. 海洋科学集刊, 11: 91-163 (in Chinese).
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}