
Journal of Tropical Oceanography >
Intracellular and extracellular metabolites analysis and key metabolite screening on the Bacillus paranthracis SG49
Copy editor: SUN Cuici
Received date: 2024-03-23
Revised date: 2024-04-03
Online published: 2024-04-28
Supported by
NSFC-Shandong Joint Fund(U2106208)
National Natural Science Foundation of China(41976147)
As a result of climate change and human intervention, jellyfish outbreaks have become a serious ecological disaster that threatens coastal economies and marine ecosystems. Globally, there is an urgent need to prevent jellyfish blooms. Microorganisms play an important role in the growth and development of marine invertebrates. Co-culture experiments revealed that Bacillus paranthracis SG49 inhibits the settlement and metamorphosis of Aurelia coerulea planula larvae. The key metabolic pathways and mechanisms behind this inhibition, however, require further investigation. Using non-targeted metabolomics technology, the intracellular and extracellular metabolites of SG49 were detected, differences between the two groups were analyzed, and potential metabolites affecting planula larval metamorphosis were identified. Our results showed that SG49 intracellular and extracellular metabolites were significantly different. Specifically, seven substances were screened for their potential inhibitory activities, including 3-hydroxy-2-oxindole, kanamycin, apramycin, streptomycin, streptomycin sulfate, gallic acid, and coniferyl alcohol. Bacterial biofilms and microorganism growth can be inhibited by these metabolites. Our findings provide a theoretical basis and strain resources to prevent jellyfish outbreaks in the future.
LIU Shuai , LIU Xuerui , ZHANG Rui , GUO Xiangrui , YU Zhen , SUN Hao , ZHANG Yanying . Intracellular and extracellular metabolites analysis and key metabolite screening on the Bacillus paranthracis SG49[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 122 -132 . DOI: 10.11978/2024068
表1 菌株SG49胞内和胞外代谢物丰度Tab. 1 The abundance of intracellular and extracellular metabolites from SG49 |
| 离子模式 | 所有峰值 | 已鉴定代谢物 | Library数据库代谢物 | KEGG数据库代谢物 |
|---|---|---|---|---|
| 阳离子 | 5618 | 1134 | 1004 | 497 |
| 阴离子 | 8835 | 927 | 895 | 454 |
图2 代谢物数据质量分析a. QC样本评估曲线, 虚线表示预处理前数据, 实线表示预处理后数据; b. 样本相关性热图 Fig. 2 The data quality analysis of all metabolites. (a) The quality evaluation curve of QC samples. Dashed lines indicate pre-pretreatment data and solid lines represent post-pretreatment data. (b) The heat map of sample correlations |
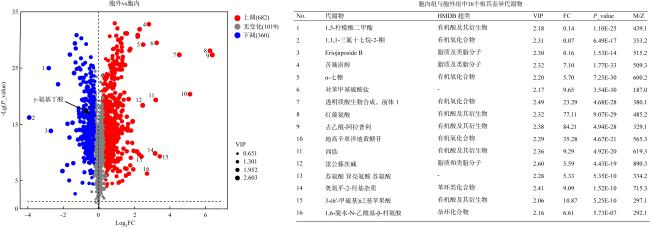
图3 菌株SG49胞内外代谢物OPLS-DA及置换检验图a. 阳离子模式下胞内组和胞外组的OPLA-DA分析; b. 阴离子模式下胞内组和胞外组的OPLA-DA分析; c. 阳离子模式下OPLA-DA分析的置换检验; d. 阴离子模式下OPLA-DA分析的置换检验 Fig. 3 The OPLS-DA plot of metabolites in SG49 intracellular and extracellular groups. (a) OPLA-DA analysis of intracellular and extracellular groups in positive ion mode; (b) OPLA-DA analysis of intracellular and extracellular groups in negative ion mode; (c) permutation testing for OPLA-DA analysis in positive ion mode; (d) permutation testing for OPLA-DA analysis in negative ion mode |
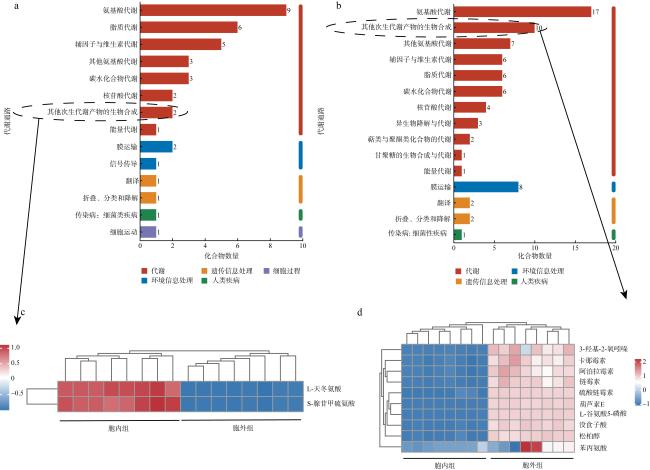
图4 代谢物注释及富集分析a. 胞内超类水平上HMDB注释的特有代谢物; b. 胞内特有代谢物KEGG通路富集分析图; c. 胞外超类水平上HMDB注释的特有代谢物; d. 胞外特有代谢物KEGG通路富集分析图; e. 胞外和胞内的样本比较Venn分析图 Fig. 4 The annotation and enrichment analysis of metabolites. (a) Unique metabolites of intracellular group at superclass level of HMDB database; (b) enrichment analysis diagram of KEGG pathway of unique metabolites in intracellular group; (c) unique metabolites of extracellular group at superclass level of HMDB database; (d) enrichment analysis diagram of KEGG pathway of unique metabolites in extracellular groups; (e) Venn analysis chart comparing samples from intracellular and extracellular groups |
图6 胞内外特有代谢物的KEGG功能通路注释a. 胞内组特有代谢物的KEGG功能通路; b. 胞外组特有代谢物的KEGG功能通路; c. 胞内组中其他次生代谢物的生物合成组物质的丰度变化; d. 胞外组中其他次生代谢物的生物合成组物质的丰度变化 Fig. 6 KEGG functional pathway of intracellular and extracellular unique metabolites. (a) KEGG pathway of unique metabolites in intracellular group; (b) KEGG pathway of unique metabolites in extracellular group; (c) the abundance of other secondary metabolite biosynthesis in intracellular group; (d) the abundance of other secondary metabolite biosynthesis in extracellular group |
| [1] |
黄文明, 尹梦丽, 陈煜, 等, 2024. 一株高效同化氨氮霉菌的筛选及其代谢组分析[J]. 微生物学报, 64(1): 161-173.
|
| [2] |
刘佳慧, 吕红, 林娟, 2023. 一株产低温碱性淀粉酶蕈状芽孢杆菌的分离筛选和发酵优化[J]. 四川大学学报(自然科学版), 60(2): 026001.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}