
Journal of Tropical Oceanography >
Study on the annual growth and early development of sexual reproduction of Sargassum wightii
Copy editor: SUN Cuici
Received date: 2024-03-23
Revised date: 2024-04-15
Online published: 2024-05-17
Supported by
National Key Research and Development Plan of China(2022YFD2401302)
Basic and Applied Basic Research Foundation of Guangdong Province, China(2022A1515010590)
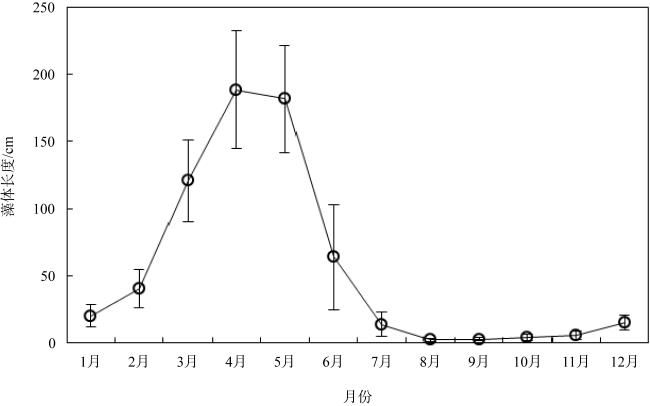
Sargassum wightii is a common species in the Weizhou Island of Guangxi, Xuwen of Guangdong and other sea areas, and is an economic macroalgae. Understanding the annual growth and early development of macroalgae can predict the mature time of thalli and provide scientific basis for artificial seedling and cultivation of macroalgae. This paper reports the annual growth characteristics, the effect of temperature on ovulation and the early developmental process of sexual reproduction of S. wightii in the sea area of Xuwen, Guangdong. The results show that the annual growth of S. wightii can be divided into 4 periods: fast growing period, mature period, decline period and resting period. The average length of thalli reaches its maximum [(188.32±43.77) cm] in early April, exceeding 180 cm in both April and May, and less than 5 cm in late July to early November. The male and female reproductive parts of S. wightii are in the same receptacle, and the egg cell development belongs to 8-nucleui type. The culture temperature is very important to the ovulation process. Ovulation could not be achieved at 15, 27 and 30 ℃, but could be achieved at 18~24℃. The ovulation time is shortest at 24 ℃, only 3 d. The zygotes develop into germlings with rhizoid buds at 24 ℃ for about 24h, and the seedlings develop a fourth leaflet after 30 d of culture. The results of this study provide basic information for the biology, ecology, artificial seedling and cultivation of S.wightii.
ZENG Zhaojun , SUN Liwei , XIE Enyi . Study on the annual growth and early development of sexual reproduction of Sargassum wightii[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 44 -52 . DOI: 10.11978/2024069
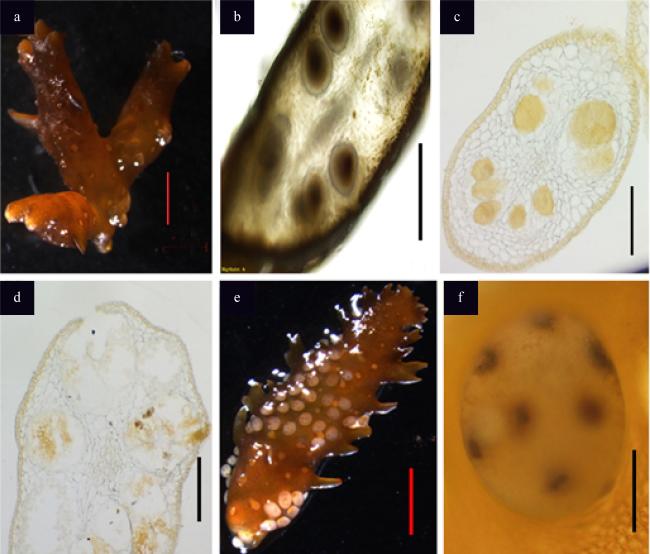
图2 成熟的围氏马尾藻形态a. 成熟藻体; b. 带有肉眼可见生殖托的藻体分枝; c. 生殖托; d. 叶片; e. 气囊; 图中比例尺长度分别为5cm (a)、2cm (b、d)、2mm (c)、1cm (e) Fig. 2 Morphology of mature S. wightii. (a) The matured thalli; (b) branchlets with visible receptacles; (c) receptacles; (d) leaves; (e) vesicles; bars=5 cm (a), 2 cm (b, d), 2 mm (c), 1 cm (e) |
图3 围氏马尾藻成熟生殖托的内部结构及卵细胞的排放a. 成熟的生殖托; b. 生殖托的活体切片; c. 生殖托的石蜡切片(雌性部分); d. 生殖托的石蜡切片(雄性部分); e. 卵细胞的排放; f. 卵细胞刚排放出来便为8核结构; 图中比例尺长度为2mm (a)、1mm (b—d)、2mm (e)、100μm (f) Fig. 3 Internal structure of receptacles and release of the eggs of S. wightii. (a) Mature receptacle; (b) vivisection of the receptacle; (c) paraffin section of the receptacle (female part); (d) paraffin section of the receptacle (male part); (e) release of the eggs; f. eggs containing eight nuclei; bars = 2 mm (a), 1 mm (b−d), 2 mm (e), 100 μm (f) |
表1 不同培养温度下生殖托的排卵时间Tab.1 Ovulation time of eggs at different temperature |
| 培养温度/℃ | 排卵时间(均值±标准差)/d |
|---|---|
| 15 | 未见排卵 |
| 18 | 15 ± 1.0 |
| 21 | 10 ± 0.5 |
| 24 | 3 ± 0.5 |
| 27 | 腐烂未排卵 |
| 30 | 腐烂未排卵 |
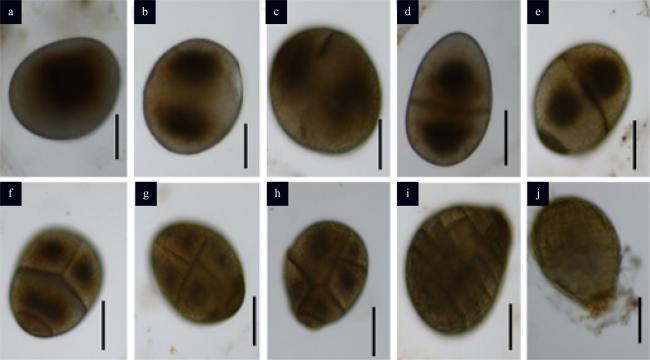
图4 围氏马尾藻受精卵的发育a. 含有一个大核的受精卵; b. 受精卵细胞核的分裂; c、d. 受精卵细胞的第一次分裂; e. 幼孢子体第二次分裂以及假根源细胞的形成; f—i. 经过几次有分裂后形成地雷状多细胞结构; j. 发育出假根的孢子体; 图中比例尺为100μm Fig. 4 Development of S. wightii zygote. (a) A zygote with a central large nucleolus; (b) the first division of large central nucleolus; (c, d) the first division of zygote across its longer axis; (e) the second division and the original cell of rhizoids; (f−i) the landmine-like germling were formed after several divisions; (j) germling with rhizoid buds; bars = 100 μm |
图5 围氏马尾藻幼孢子体的发育和生长a. 培养24h后的幼孢子体; b. 培养36h后的幼孢子体; c. 培养48h后的幼孢子体; d. 培养4d后的幼孢子体; e. 第一片叶片的发育形成; f. 培养8d后的幼孢子体; g. 培养10d后的幼孢子体; h. 培养15d后的幼孢子体; i. 培养20d后的幼孢子体; j. 培养30 d后的幼孢子体; 图中比例尺分别为200μm (a—d)、500μm (e—h)、1mm (i—j) Fig. 5 Development and growth of S. wightii germlings. (a) Germlings after being cultivated for 24h; (b) germlings after being cultivated for 36h; (c−d) germlings after 2 and 4d of cultivation; (e) the first leaflet; (f) germlings after 8d of cultivation; (g) germlings after 10d of cultivation; (h−j) germlings after 15, 20, and 30d of cultivation with aeration; bars = 200 μm (a−d), 500 μm (e−h), and 1mm (i−j) |
| [1] |
曹淑青, 张泽宇, 王国书, 等, 2008. 海黍子室内人工育苗技术的研究[J]. 大连水产学院学报, 23(5): 359-363.
|
| [2] |
郭伟, 邓巍, 燕雪飞, 等, 2010. 植物生殖分配影响因素的研究进展[J]. 东北农业大学学报, 41(9): 150-156.
|
| [3] |
贾柽, 杨彬, 谢恩义, 2012. 莫氏马尾藻繁殖生物学初步研究[J]. 水产科学, 31(10): 616-619.
|
| [4] |
李大鹏, 李慧, 张桂军, 等, 2018. 铜藻繁殖发育过程的研究[J]. 海洋科学, 42(5): 40-45.
|
| [5] |
李伟新, 朱仲嘉, 刘凤贤, 1982. 海藻学概论[M]. 上海: 上海科学技术出版社: 224-225 (in Chinese).
|
| [6] |
林立东, 尚天歌, 张甜甜, 等, 2020. 褐藻羊栖菜繁殖生物学特征和生活史流程的补充研究[J]. 水产学报, 44(4): 581-595.
|
| [7] |
毛晓霞, 2013. 石蜡切片制作方法的改良[J]. 安徽农学通报, 19(8): 15-16, 39.
|
| [8] |
潘金华, 张全胜, 许博, 2007. 鼠尾藻有性繁殖和幼孢子体发育的形态学观察[J]. 水产科学, 26(11): 589-592.
|
| [9] |
彭燕, 陈思, 肖生鸿, 等, 2017. 围氏马尾藻的营养化学成分及评价[J]. 天然产物研究与开发, 29(2): 249-254.
|
| [10] |
曲元凯, 2015. 三种马尾藻的生长繁殖和人工藻场的构建[D]. 湛江: 广东海洋大学.
|
| [11] |
曲元凯, 谢恩义, 孙立伟, 等, 2015. 3种马尾藻周年生长和成熟时间以及形态学研究[J]. 安徽农业科学, (23): 136-139, 231.
|
| [12] |
阮积惠, 徐礼根, 2001. 羊栖菜Sargassum fusiforme Setch繁殖与发育生物学的初步研究[J]. 浙江大学学报(理学版), 28(3): 315-320.
|
| [13] |
孙建璋, 陈万东, 庄定根, 等, 2008a. 中国南麂列岛铜藻Sargassum horneri实地生态学的初步研究[J]. 南方水产, 4(3): 58-63.
|
| [14] |
孙建璋, 庄定根, 陈万东, 等, 2008b. 铜藻Sargassum horneri繁殖生物学及种苗培育研究[J]. 南方水产, 4(2): 6-14.
|
| [15] |
孙立伟, 谢恩义, 曲元凯, 2015. 亨氏马尾藻的有性生殖观察[J]. 南方水产科学, 11(1): 62-67.
|
| [16] |
孙立伟, 2016. 围氏马尾藻繁殖生物学的初步研究[D]. 湛江: 广东海洋大学.
|
| [17] |
王飞久, 孙修涛, 李锋, 2006. 鼠尾藻的有性繁殖过程和幼苗培育技术研究[J]. 海洋水产研究, 27(5): 1-6.
|
| [18] |
魏晓娇, 2012. 温度和光强对带形蜈蚣藻(Grateloupia turuturu)丝状体发育的影响及白氏鹿角菜(Silvetia babingtonii)的早期发育[D]. 青岛: 中国海洋大学.
|
| [19] |
吴海一, 丁刚, 刘玮, 等, 2019. 经济马尾藻类海藻苗种规模化繁育及人工增殖技术[J]. 中国科技成果, 20(2): 58-60 (in Chinese).
|
| [20] |
许博, 2009. 鼠尾藻(Sargassum thunbergii)繁殖生态学研究[D]. 青岛: 中国海洋大学.
|
| [21] |
曾呈奎, 2000. 中国海藻志: 褐藻门[M]. 北京: 科学出版社: 56-58 (in Chinese).
|
| [22] |
张才学, 周伟男, 孙省利, 等, 2020. 硇洲岛大型海藻群落的季节演替[J]. 热带海洋学报, 39(1): 74-84.
|
| [23] |
章守宇, 刘书荣, 周曦杰, 等, 2019. 大型海藻生境的生态功能及其在海洋牧场应用中的探讨[J]. 水产学报, 43(9): 2004-2014.
|
| [24] |
邹定辉, 高坤山, 2010. 羊栖菜离体生殖托低温超低温的保存[J]. 水产学报, 34(6): 935-941.
|
| [25] |
猪野俊平, 1947, [2024-03-23]. 海藻の發生[M/OL]. 东京: 国立国会図書館, 67-81. https://ndlsearch.ndl.go.jp/books/R100000002-I030177533 (in Japanese).
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}