
Journal of Tropical Oceanography >
Molecular cloning and functional study of cyclic GMP-AMP synthase from Crassostrea gigas
Copy editor: SUN Cuici
Received date: 2024-03-22
Revised date: 2024-04-10
Online published: 2024-05-21
Supported by
National Key Research and Development Program of China(2022YFD2400301)
National Natural Science Foundation of China(32073002)
National Natural Science Foundation of China(U22A20533)
9th Young Elite Scientists Sponsorship Program(2023QNRC001)
Science and Technology Planning Project of Guangdong Province, China(2023A04J0096)
Science and Technology Planning Project of Guangdong Province, China(2024A04J6278)
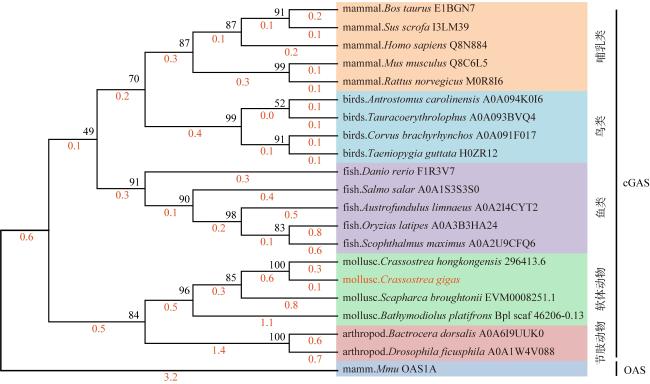
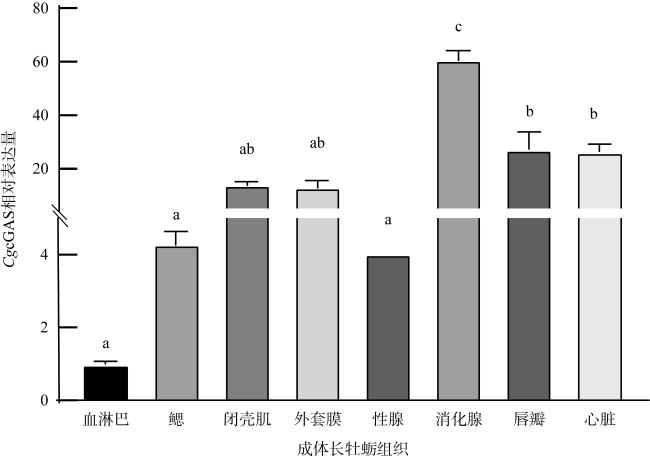
Cyclic GMP-AMP synthase (cGAS) is a critical intracellular sensor that can recognize abnormally located DNA in the cytoplasm and trigger immune responses. To elucidate the critical role of cGAS in the regulation of innate immunity in mollusks, we successfully cloned and analyzed Crassostrea gigas cGAS (CgcGAS). The open reading frame (ORF) of CgcGAS was 1623bp and encoded 540 amino acids with a theoretical molecular weight of 62.3 kDa and a conserved Mab21 domain. Phylogenetic analysis confirmed that CgcGAS was a member of the molluscan cGAS family. Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) results revealed widespread expression of CgcGAS in various tissues, with the highest relative expression in the digestive glands. Subsequently, subcellular localization experiments showed that CgcGAS was observed in both the nucleus and cytoplasm, with a predominant nuclear localization, suggesting that CgcGAS may have played a role in DNA sensing in the nucleus and DNA binding and signaling in the cytoplasm. Furthermore, dual-luciferase reporter gene assays and RNA interference experiments revealed that CgcGAS could activate the NF-κB and ISRE signaling pathways, as well as the expression of downstream inflammation-related factors, such as virus inhibitory protein endoplasmic reticulum-associated interferon-inducible (viperin), tumor necrosis factor (TNF), interleukin-17 (IL-17), and the transcription factor interferon regulatory factor 2/8 (IRF2/8). In conclusion, CgcGAS played a critical role in the signal transduction process of innate immune responses in Crassostrea gigas.
Key words: Crassostrea gigas; innate immunity; cyclic GMP-AMP synthase; gene cloning; function
BAI Jing , MAO Fan , LIU Kelin , SONG Jingchen , YU Ziniu , ZHANG Yang . Molecular cloning and functional study of cyclic GMP-AMP synthase from Crassostrea gigas[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 24 -34 . DOI: 10.11978/2024067
表1 引物信息Tab. 1 Sequences of designed primers used in this study |
| 引物名称 | 序列(5′-3′) | 用途 |
|---|---|---|
| cGAS-F1 | ATGGTAATCAAATGTCCTAATTGTG | ORF克隆 |
| cGAS-R1 | TTATTGTAAGAGACCCTCTAGTTCC | ORF克隆 |
| pEGFP-N1-cGAS-F | TCAGATCTCGAGCTCAAGCTTGCCACCATGGTAATCAAATGTCC | pEGFP-N1-cGAS重组质粒 |
| pEGFP-N1-cGAS-R | ATGGTGGCGACCGGTGGATCCGATTGTAAGAGACCCTCTAGTTCCCT | pEGFP-N1-cGAS重组质粒 |
| PCDNA3.1/V5-His-cGAS-F | GCACAGTGGCGGCCGCTCGAGATGGTAATCAAATGTCCTAATTGTGAC | pcDNA3.1/V5-His-cGAS重组质粒 |
| PCDNA3.1/V5-His-cGAS-R | AGGCTTACCTTCGAAGGGCCCTTGTAAGAGACCCTCTAGTTCCCTG | pcDNA3.1/V5-His-cGAS重组质粒 |
| dscGAS-F | CGACAAAACAAAGATCGACTACAAC | cGAS-RNAi |
| dscGAS-R | CAGTCTGGATTTCTCTTGCATCCTT | cGAS-RNAi |
| dscGAS-F-T7 | TAATACGACTCACTATAGGCGACAAAACAAAGATCGACTACAAC | cGAS-RNAi |
| dscGAS-R-T7 | TAATACGACTCACTATAGGCAGTCTGGATTTCTCTTGCATCCTT | cGAS-RNAi |
| dsGFP-F | GCAAGGGCGAGGAGCTGTTCACCGG | GFP-RNAi |
| dsGFP-R | TTGCCGTCCTCCTTGAAGTCGATGC | GFP-RNAi |
| dsGFP-F-T7 | TAATACGACTCACTATAGGGCAAGGGCGAGGAGCTGTTCACCGG | GFP-RNAi |
| dsGFP-R-T7 | TAATACGACTCACTATAGGTTGCCGTCCTCCTTGAAGTCGATGC | GFP-RNAi |
| cGAS-F2 | GGAAAGACGACAGGGACGG | qRT-PCR |
| cGAS-R2 | TGTCTGGAGAACCCCTTTGG | qRT-PCR |
| viperin-F | CTGAAACCCATCAGTGTCAACTACC | qRT-PCR |
| viperin-R | GACAATGAAGGGCTCGCCAC | qRT-PCR |
| IRF2-F | ACTTCCGCTGTGCCCTGAAT | qRT-PCR |
| IRF2-R | TATGACCTTTGGCACTGTCGTTC | qRT-PCR |
| IL-17-F | AAACATGCTGGAATACCTCGAGT | qRT-PCR |
| IL-17-R | GTGGGACGCTACGAGGAAATA | qRT-PCR |
| TNF-F | GTATTACTGAGGAATGGCGTGAAAC | qRT-PCR |
| TNF-R | CAAAATCCCTGTCACTACAACCG | qRT-PCR |
| IRF8-F | GGACAGCGGTCAGACACGAC | qRT-PCR |
| IRF8-R | CCTTGAATATTGTGGAGTCTGCCT | qRT-PCR |
| β-actin-F | GGATTGGCGTGGTGGTAGAG | qRT-PCR |
| β-actin-R | GTATGATGCCCCTTTGTTGAGTC | qRT-PCR |
注: “F”表示上游引物, “R”表示下游引物 |
表2 双荧光素报告基因质粒转染体系Tab. 2 Plasmids transfection in dual luciferase reporter assay |
| 组别 | pNF-κB-luc/ISRE-luc/ng | 目的质粒/ng | 对照空质粒/ng | 内参质粒RT-TK/ng |
|---|---|---|---|---|
| 对照组 | 200 | 0 | 400 | 20 |
| 实验组1 | 200 | 100 | 300 | 20 |
| 实验组2 | 200 | 200 | 200 | 20 |
| 实验组3 | 200 | 400 | 0 | 20 |
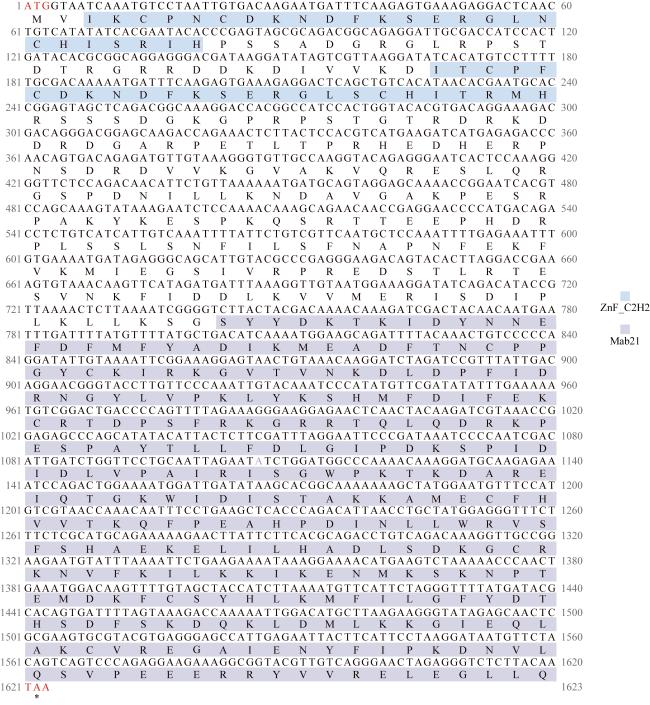
图1 长牡蛎cGAS开放阅读框序列全长和推导的氨基酸序列锌指结构域用蓝色方框标出, Mab21结构域用紫色方框标出; *表示蛋白翻译终止, 起始密码子ATG和终止密码子TAA为红色字体 Fig. 1 Full length of the Crassostrea gigas cGAS open reading frame sequence and deduced amino acid sequence. The zinc finger structure is marked with a blue box and the Mab21 structural domain is marked with a purple box. * indicates protein translation termination, with initiation codon ATG and termination codon TAA in red font |
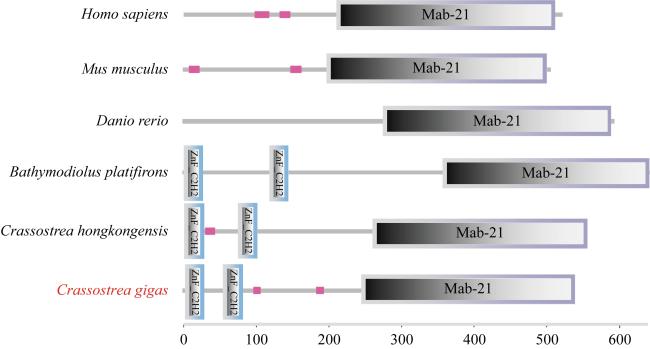
图2 cGAS在不同物种中的结构图解人(Homo sapiens)、小鼠(Mus musculus)、斑马鱼(Danio rerio)、深海偏顶蛤(Bathymodiolus platifron)、香港牡蛎(Crassostrea hongkongensis)和长牡蛎(Crassostrea gigas) Fig. 2 Schematic of cGAS structures in different species |
图3 长牡蛎与其他物种cGAS氨基酸序列比对图中NTase活性位点残基以红点标记, 锌带结构域以方框标出, “---” 表示氨基酸缺失; “*” “:” 和“.”分别表示相同的、高度保守的和不太保守的氨基酸残基 Fig. 3 Multiple alignment of cGAS amino acid sequences between Crassostrea gigas and other species. NTase catalytical sites are marked with red dots, and the zinc-ribbon domain is indicated by a box, with “---” representing an amino acid deletion. “*”, “:” and “.” present identical, highly conserved, and less conserved amino acid residues, respectively |
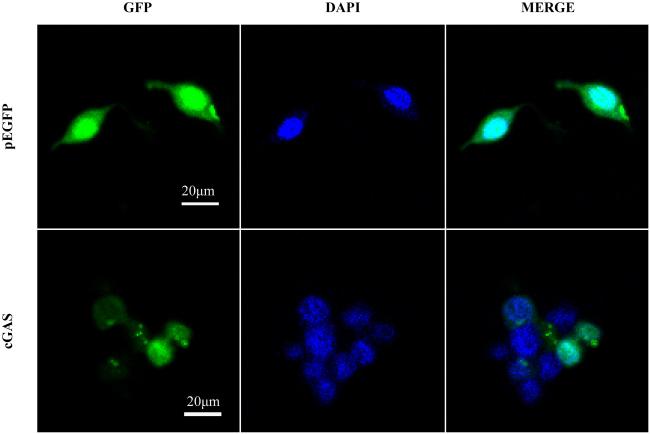
图6 长牡蛎cGAS的亚细胞定位左侧为pEGFP与cGAS荧光融合蛋白, 中间为DAPI染色后的细胞核, 右侧为合并图像。40倍镜下HEK293T细胞显微观察结果 Fig. 6 Subcellular localization of cGAS in Crassostrea gigas. Shown on the left is the fluorescent fusion protein of pEGFP and cGAS; shown in the middle is the nucleus after DAPI staining; and shown on the right is the combined image. The microscopic observation results of HEK293T cells at 40 magnifications |
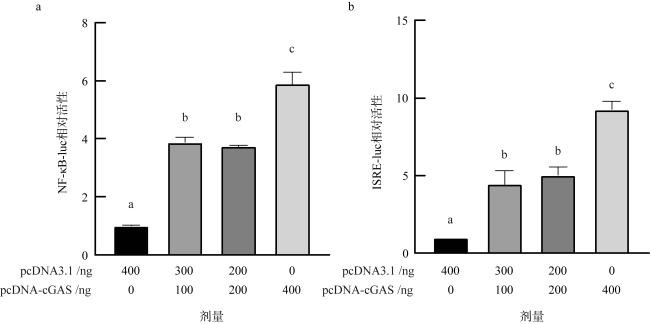
图7 长牡蛎中cGAS对NF-κB(a)和ISRE-luc(b)的激活效果数据均以平均值±SEM表示, n=3, 显著性差异(P<0.05)由不同字母a—c表示 Fig. 7 The activating effect of cGAS on NF-κB (a) and ISRE-luc (b) in Crassostrea gigas. All data are expressed as mean±SEM, n = 3. Significant differences (P< 0.05) are identified by different letters, a-c |
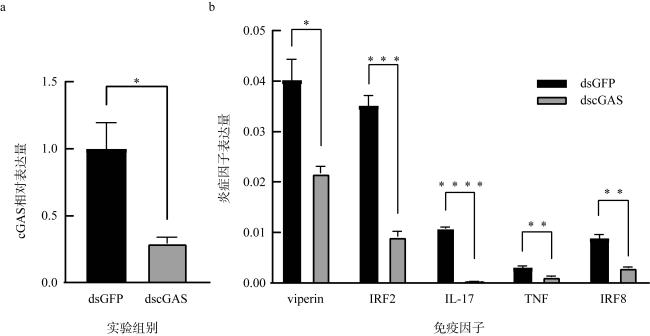
图8 长牡蛎cGAS敲降后下游因子表达情况a. 荧光定量PCR检测敲降效果; b. cGAS敲降后下游免疫因子表达量; 数据均以平均值±SEM表示, n=3, *表示 P<0.05, **表示P<0.01, ***表示P<0.001, ****表示P<0.0001 Fig. 8 Expression of downstream factors after cGAS knockdown in Crassostrea gigas; (a) fluorescence quantitative PCR was used to detect the knockdown effect; (b) expression of downstream immune factors after cGAS knockdown. All data are presented as mean ± SEM, with significance denoted as * for P<0.05, ** for P<0.01, *** for P<0.001, and **** for P<0.0001 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}