
Journal of Tropical Oceanography >
Observation on the asexual reproduction of Phyllorhiza sp.
Copy editor: SUN Cuici
Received date: 2024-03-19
Revised date: 2024-04-15
Online published: 2024-05-23
Supported by
Guided Project of Fujian Provincial Department of Science and Technology(2022N0014)
The Regional Demonstration Project of the 13th Five-Year Plan of Marine Economy Innovation & Development in Xiamen(16PZY002SF18)
In this experiment, the conditions for amplifying polyp populations and strobilation of Phyllorhiza sp. were studied. The asexual reproductive process and the metamorphosis of ephyra into medusa were observed and described. Phyllorhiza sp. is characterized by two asexual reproduction modes: swimming buds and strobilation. Polyps can be permanently stored in laboratory. Both the producing swimming buds and strobilation can be observed at 20~25°C. It takes 10~12 days for metamorphosis to mature polyps after swimming buds attachment. It takes 2~3 days to produce swimming buds for polyp asexual reproduction. Monodisc is the strobilation mode of Phyllorhiza sp. and it takes 4~5 days before the ephyra gets rid of the residual polyp. The edge of the central disc of strobila keeps blue-purple from the formation of rhopalia to the disappearing of polyp tentacles. It takes 12 days for ephyra to develop into juvenile medusa under 25°C, 29‰~32‰ (salinity).
Key words: Phyllorhiza sp.; polyp; ephyra; asexual reproduction; morphology
HAO Yanxia , ZHAO Yuxuan , DENG Liqiu , ZHANG Ruixue , WANG Shuhong . Observation on the asexual reproduction of Phyllorhiza sp.[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 35 -43 . DOI: 10.11978/2024062
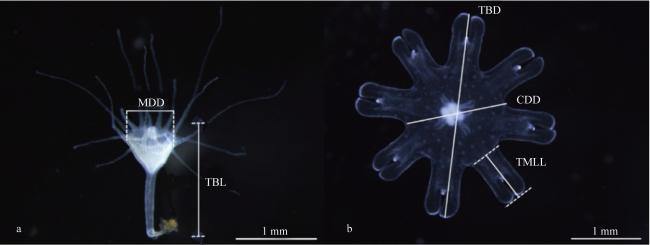
图1 蓝环水母(Phyllorhiza sp.)水螅体(a)和碟状体(b)形态指标测量方式示意图TBL: 体长; MDD: 水螅体口盘直径; TBD: 碟状体直径; CDD: 中心口盘直径; TMLL: 缘瓣长度 Fig. 1 Schematic diagram of the measurement method of morphological parameters of the polyp and ephyra in Phyllorhiza sp.; (a) the polyp; (b) the ephyra; TBL, total body length; MDD, mouth disc diameter; TBD, total body diameter; CDD, central disc diameter; TMLL, total marginal lappet length |
表1 蓝环水母(Phyllorhiza sp.)自由游泳胞芽附着发育成水螅体的过程(n=10)Tab. 1 Metamorphosis from planuloid to polyp of Phyllorhiza sp. |
| 发育过程 | 时间/d | 长(胞芽或水螅体TBL)/(μm) | 宽(胞芽或水螅体MDD/(μm) |
|---|---|---|---|
| 刚附着的胞芽 | 0 | 350±29 | 232±29 |
| 8触手水螅体 | 1 | 550±119 | 315±35 |
| 10触手水螅体 | 1—2 | 650±158 | 359±35 |
| 12触手水螅体 | 2—4 | 746±231 | 363±38 |
| 14触手水螅体 | 3—6 | 821±169 | 379±44 |
| 16触手水螅体 | 4—8 | 1025±161 | 405±40 |
| 成熟水螅体 | 10—12 | 1925±279 | 674±89 |
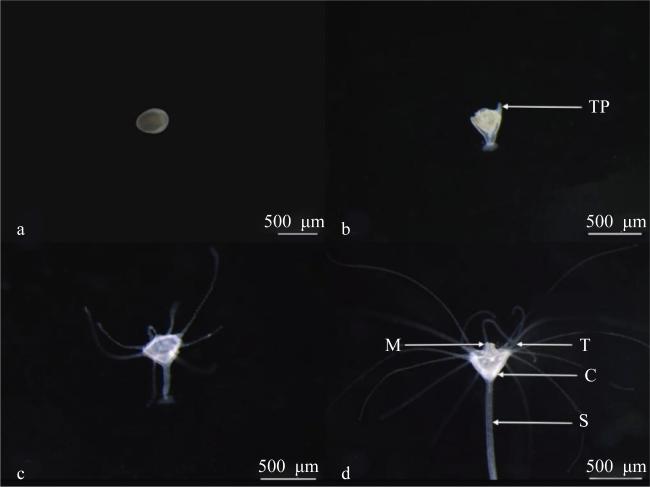
图3 蓝环水母(Phyllorhiza sp.)自由游泳胞芽附着发育成水螅体的过程a. 胞芽; b. 具4个触手原基的水螅体; c. 8触手水螅体; d. 16触手水螅体 Fig. 3 Metamorphosis from planuloid to polyp of Phyllorhiza sp.; (a) planuloid; (b) polyp with four tentacle primordias; (c) polyp with eight tentacles; (d) polyp with sixteen tentacles; TP, tentacle primordia; T, tentacle; M, manubrium; C, calyx; S, stalk |
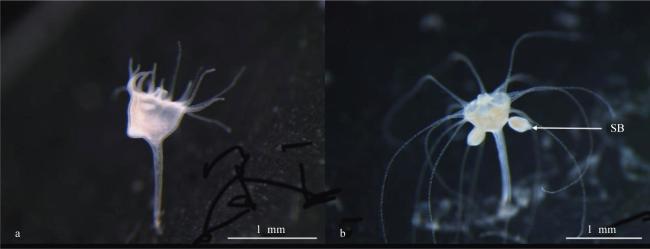
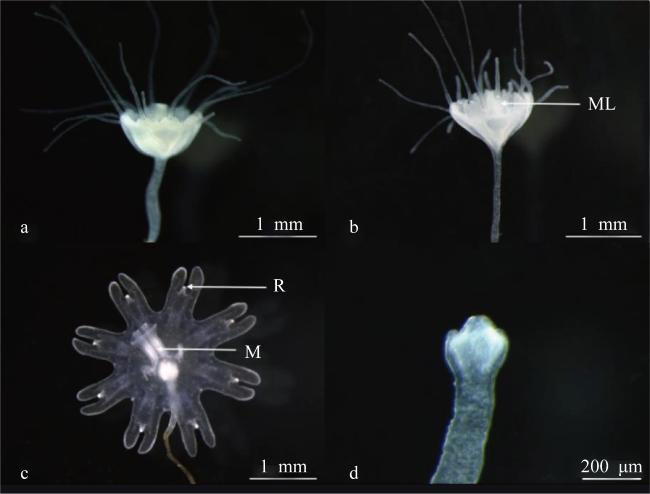
图4 蓝环水母(Phyllorhiza sp.)水螅体的横裂生殖a. 触手基部膨大; b. 触手收缩; c. 碟状体即将释放; d. 出碟后的水螅体; ML: 缘叶; R: 感觉器; M: 口 Fig. 4 The development process of strobilation in Phyllorhiza sp.; (a) Basal expansion of tentacles; (b) Tentacle contraction; (c) the releasing of ephyra; (d) Polyp after ephyra released; ML, marginal lobes; R, rhopalium; M, manubrium |
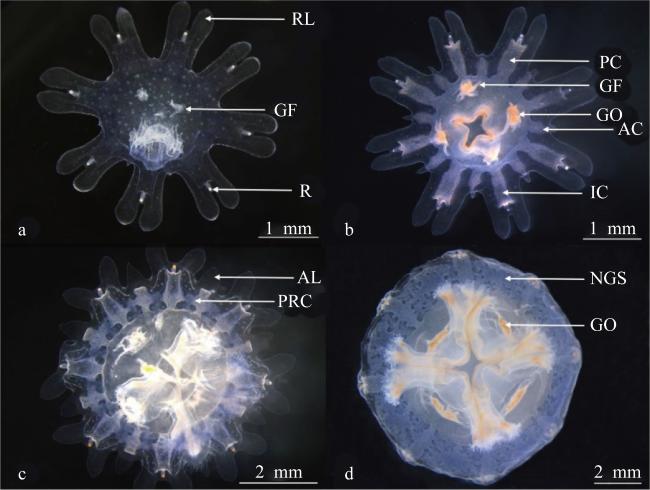
图5 蓝环水母(Phyllorhiza sp.)碟状体发育过程a. 刚脱落的碟状体; b. 投喂4h后的碟状体; c. 最初环管形成; d. 网状循环系统形成 Fig. 5 The development process of the ephyra in Phyllorhiza sp.; (a) newly released ephyra; (b) ephyra after 4 hours of feeding; (c) primary ring canal formation; (d) net-like gastric system formation; RL, rhopalar lappet; GF, gastric filament; R, rhopalium; PC, perradial canal; GO, gastric ostia; AC, adradial canal; IC, interradial canal; AL, adradial lappet; PRC, primary ring canal; NGS, net-like gastric system |
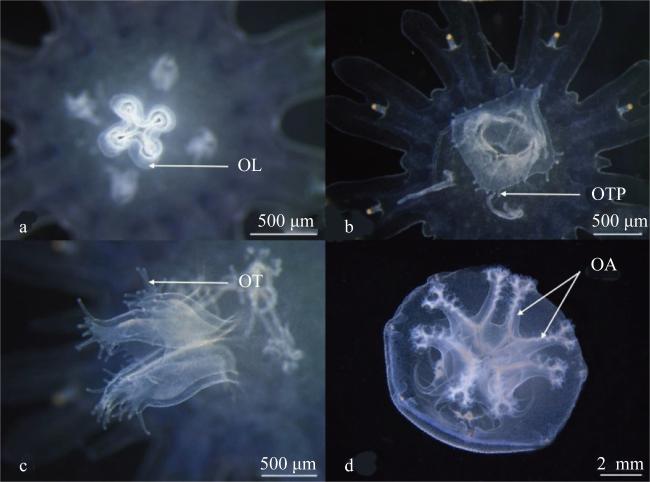
图6 蓝环水母(Phyllorhiza sp.)口的发育过程a. 刚脱落碟状体的口唇; b. 形成口触指原基; c. 口唇出现凹槽; d. 口腕形成; OL: 口唇; OTP: 口触指原基; OT: 口触指; OA: 口腕 Fig. 6 Oral lips development process of Phyllorhiza sp.; (a) oral lips view of newly released ephyra; (b) oral tentacle primordia formed; (c) oral lips appeared groove; (d) oral arms formation; OL, oral lips; OTP, oral tentacle primordia; OT, oral tentacle; OA, oral arms |
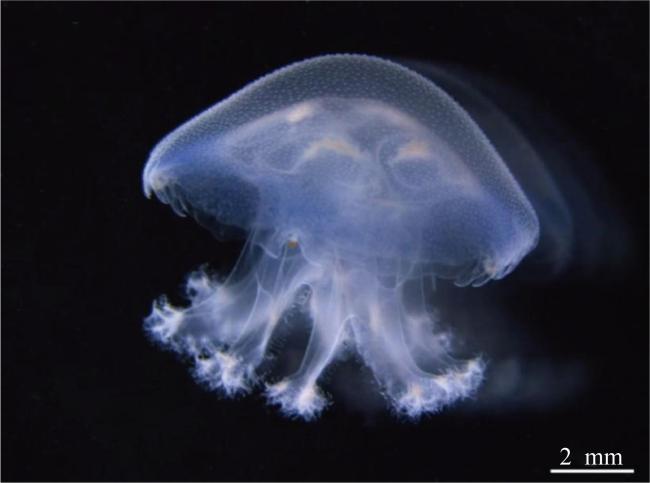
表2 蓝环水母(Phyllorhiza sp.)碟状体发育成水母幼体的过程(n=10)Tab. 2 The process from ephyra to juvenile medusa of Phyllorhiza sp. |
| 发育过程 | 时间/d | TBD/mm |
|---|---|---|
| 刚脱落的碟状体 | 0 | 3.69±0.42 |
| 循环系统最初环管形成 | 5—7 | 6.83±0.77 |
| 网状循环系统 | 8—10 | 9.47±1.08 |
| 形成8个口腕 | 11—13 | 10.88±1.14 |
| [1] |
董婧, 王彬, 刘春洋, 2006. 白色霞水母各发育阶段的形态[J]. 水产学报, 30(6): 761-766.
|
| [2] |
付瑶, 董志军, 刘东艳, 2012. 海月水母无性生殖的影响因子[J]. 生态科学, 31(3): 335-339.
|
| [3] |
林政飞, 王淑红, 2019. 钵水母水螅体横裂诱发条件及调控机制研究进展[J]. 应用生态学报, 30(3): 1057-1066.
|
| [4] |
刘春胜, 庄志猛, 陈四清, 等, 2014. 海蜇生活史的补充研究[J]. 海洋科学, 38(12): 16-21.
|
| [5] |
马永全, 于新, 黄小红, 2009. 海蜇的药用与食用价值研究进展[J]. 广东农业科学, (9): 153-156, 165.
|
| [6] |
孙明, 段妍, 柴雨, 等, 2020. 海月水母螅状体无性繁殖方式观察及一种新发现报道[J]. 水产学报, 44(12): 2007-2016.
|
| [7] |
杨翠华, 王玮, 王文章, 2011. 安朵仙水母生活史的实验观察[J]. 渔业科学进展, 32(5): 135-140.
|
| [8] |
杨文浩, 王淑红, 林子路, 等, 2022. 巴布亚硝水母无性生殖过程及形态变化[J]. 动物学杂志, 57(3): 321-335.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}