
Journal of Tropical Oceanography >
2-Hydroxyphenyl thiazoline derivatives and their biosynthetic gene clusters from the mangroves-derived Strepomyces ardesiacus
Copy editor: YIN Bo
Received date: 2024-04-02
Revised date: 2024-05-21
Online published: 2024-05-23
Supported by
Guangxi Science and Technology Base and Talent Special Project(Guike AD22035167)
Natural Science Foundation of Guangxi Province(2023GXNSFBA026139)
Natural Science Foundation of Guangxi Province(2023GXNSFAA026313)
Open Project of Guangdong Key Laboratory of Marine Materia Medica(LMM2022-4)
Initial Scientific Research Foundation of Introduced Doctors in 2021 of Guangxi University of Chinese Medicine(2021BS022)
Guangxi University of Chinese Medicine “Guipai Traditional Chinese Medicine Inheritance and Innovation Team” Project(2022A007)
Special Fund for Bagui Scholars of Guangxi(05019055)
Guangxi University of Traditional Chinese Medicine the third batch of “Qhuang Project” high-level talent team cultivation project(202407)
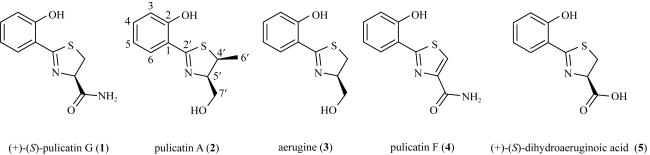
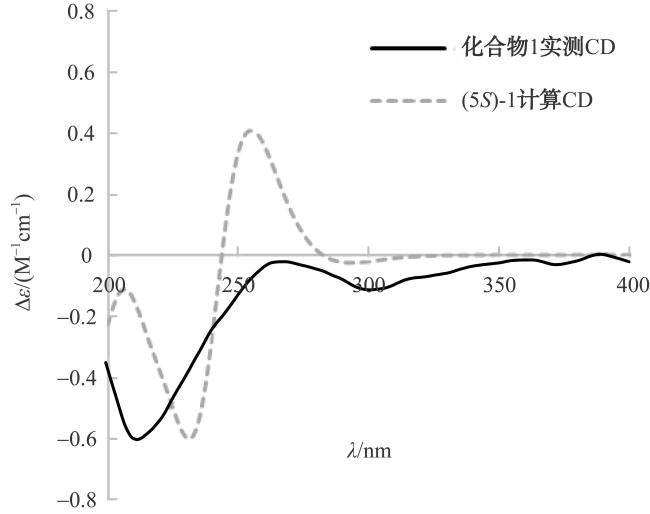
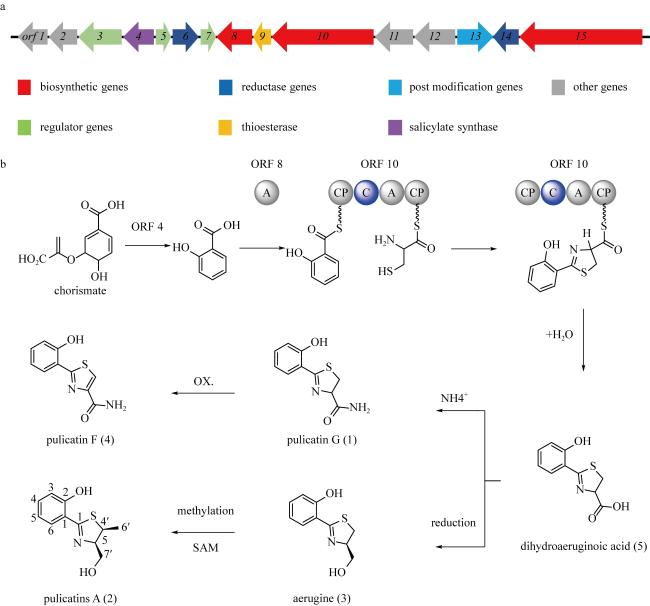
In order to identify secondary metabolites and analyze the corresponding biosynthetic gene clusters, the strain GXIMD 03502, isolated from the mangroves in Hainan, was studied. The strain was identified by comparing the 16S rDNA sequence. The secondary metabolites were separated by various chromatographic separation techniques, and their structures were determined by spectral data analysis and literature comparison. Five 2-hydroxyphenylthiazoline derivatives were isolated from this strain and identified as (+)-(S)-pulicatin G (1), pulicatin A (2), aerugine (3), pulicatin F (4), and (+)-(S)-dihydraeruginoic acid (5). At the same time, the genomic DNA of the strain was obtained by Illumina novaseq 6000 sequencing technology. The bioinformatic approach was employed to annotate the biosynthetic gene cluster and deduce the biosynthetic pathways of the compounds. The analysis results of the whole genome DNA sequence indicated that it contained 31 secondary metabolic biosynthetic gene clusters, and revealed that cluster 11 may be responsible for the biosynthesis of 2-hydroxyphenylthiazoline derivatives. This study provided a mangroves-derived S. ardesiacus GXIMD 03502 to produce 2-hydroxyphenylthiazoline derivatives, and the whole genome sequence of this strain laid a foundation for the exploration of its secondary metabolites.
LIU Sini , BAI Meng , ZHU Yiguang , LUO Xiaowei , GAO Chenghai , LIU Yonghong , XU Xinya , JIANG Xiaodong . 2-Hydroxyphenyl thiazoline derivatives and their biosynthetic gene clusters from the mangroves-derived Strepomyces ardesiacus[J]. Journal of Tropical Oceanography, 2025 , 44(2) : 64 -72 . DOI: 10.11978/2024073
表1 菌株GXIMD 03502基因组中次级代谢产物基因簇的antiSMASH分析结果Tab. 1 Secondary metabolite gene clusters of GXIMD 03502 identified by antiSMASH |
| 编号 | 类型 | 大小/kb | 相似基因簇 | 相似度/% |
|---|---|---|---|---|
| 1 | RiPP-like | 10.2 | informatipeptin | 42 |
| 2 | T3PKS | 41.2 | alkylresorcinol | 100 |
| 3 | betalactone | 27.9 | / | / |
| 4 | Terpene | 21.0 | albaflavenone | 100 |
| 5 | Phosphonate-ladderane-Ⅰ PKS | 67.2 | amycomicin | 87 |
| 6 | Terpene | 22.7 | isorenieratene | 62 |
| 7 | indole | 21.1 | 5-dimethylallylindole-3-acetonitrile | 100 |
| 8 | terpene | 23.9 | / | / |
| 9 | NRP-metallophore | 63.0 | coelibactin | 100 |
| 10 | NRP-metallophore | 58.3 | scabichelin | 100 |
| 11 | Ⅰ PKS - Ⅲ PKS-NRPS | 95.3 | thiazostatin/watasemycin | 100 |
| 12 | T3PKS | 41.1 | flaviolin | 100 |
| 13 | melanin | 10.5 | istamycin | 5 |
| 14 | NI-siderophore | 11.8 | desferrioxamin B and E | 83 |
| 15 | T1PKS-other | 60.6 | macrotermycins | 65 |
| 16 | terpene | 22.2 | geosmin | 100 |
| 17 | NRPS | 35.4 | ibomycin | 12 |
| 18 | Ⅲ PKS | 41.2 | germicidin | 100 |
| 19 | oligosaccharide | 24.9 | vancomycin glucose | 42 |
| 20 | Ⅱ PKS | 72.5 | WS-5995 | 69 |
| 21 | NI-siderophore | 11.9 | / | / |
| 22 | terpene | 26.7 | hopene | 100 |
| 23 | lanthipeptide-class-iii | 22.8 | SapB | 100 |
| 24 | ectoine | 10.4 | ectoine | 100 |
| 25 | NRPS | 25.9 | paulomycin | 13 |
| 26 | terpene | 20.9 | azinomycin B | 6 |
| 27 | T2PKS | 70.1 | spore pigment | 66 |
| 28 | NRPS | 53.2 | detoxin S1 | 33 |
| 29 | T1PKS | 38.5 | butyrolactol A | 80 |
| 30 | T1PKS-terpene | 43.3 | azalomycin F3a | 32 |
| 31 | T1PKS | 14.3 | ECO-0501 | 14 |
注: “/”表示未发现相似的基因簇 |
表2 菌株GXIMD 03502中pulicatins生物合成基因簇内各基因的功能注释Tab. 2 Deduced function of open reading frames in the pulicatin biosynthetic gene cluster from GXIMD 03502 |
| 基因 | 氨基酸长度 | 同源蛋白 | 同一性/相似性/% | 推测的基因功能 |
|---|---|---|---|---|
| orf1 | 440 | S. venezuelae ATCC10712 sven0503 (CCA53790) | 77/81 | AMP-ligase |
| orf2 | 421 | S. venezuelae ATCC10712 sven0504 (CCA53791) | 83/87 | Na+/H+ antiporter |
| orf3 | 647 | S. venezuelae ATCC10712 sven0505 (CCA53792) | 85/88 | AfsR-family regulator |
| orf4 | 455 | S. venezuelae ATCC10712 sven0506 (CCA53793) | 79/85 | Salicylate synthase |
| orf5 | 282 | S. venezuelae ATCC10712 sven0507 (CCA53794) | 85/91 | TetR-family regulator |
| orf6 | 390 | S. venezuelae ATCC10712 sven0508 (CCA53795) | 75/81 | Reductase |
| orf7 | 228 | S. venezuelae ATCC10712 sven0509 (CCA53796) | 90/94 | TetR-family regulator |
| orf8 | 532 | S. venezuelae ATCC10712 sven0510 (CCA53797) | 85/90 | Salicyl-AMP-ligase |
| orf9 | 265 | S. venezuelae ATCC10712 sven0511 (CCA53798) | 74/80 | Thioesterase |
| orf10 | 1517 | S. venezuelae ATCC10712 sven0512 (CCA53799) | 76/82 | NRPS |
| orf11 | 569 | S. venezuelae ATCC10712 sven0513 (CCA53800) | 91/95 | ABC transporter permease/ATPase |
| orf12 | 613 | S. venezuelae ATCC10712 sven0514 (CCA53801) | 89/93 | ABC transporter permease/ATPase |
| orf13 | 534 | S. venezuelae ATCC10712 sven0515 (CCA53802) | 89/94 | Radical SAM methyltransferase |
| orf14 | 383 | S. venezuelae ATCC10712 sven0516 (CCA53803) | 78/80 | Reductase |
| orf15 | 1829 | S. venezuelae ATCC10712 sven0517 (CCA53804) | 84/89 | NRPS |
图4 菌株GXIMD 03502中2-羟基苯基噻唑啉类似物的生物合成基因簇(a)和生物合成途径(b)a. 2-羟基苯基噻唑啉类似物的生物合成基因簇示意图; b. 2-羟基苯基噻唑啉类似物生物合成途径的推导。图中ORF4表示水杨酸合成酶(salicylate synthase); ORF8表示水杨酸腺苷连接酶(salicyl-AMP-ligase); ORF10表示非核糖体肽合成酶(nonribosomal peptide synthetase, NRPS); OX.表示oxidation; SAM为 S-adenosylmethionine; PCP表示肽基载体蛋白结构域(peptide carrier protein domain); A表示腺苷化结构域(adenylation domain); C表示缩合结构域(condensation domain) Fig. 4 The biosynthetic gene cluster (a) and the biosynthetic pathway (b) of pulicatins from GXIMD 03502 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}