
Journal of Tropical Oceanography >
Reproductive habit and embryonic development of the Harlequin Shrimp, Hymenocera picta (Decapoda: Hymenoceridae)
Copy editor: YIN Bo
Received date: 2024-04-16
Revised date: 2024-05-31
Online published: 2024-06-12
Supported by
Guided Project of Fujian Provincial Department of Science and Technology(2022N0014)
Regional Demonstration Project of the 13th Five-Year Plan of Marine Economy Innovation & Development in Xiamen(16PZY002SF18)
Research start-up funds of Jimei University(ZQ2021023)
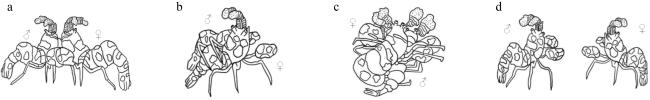
Hymenocera picta is popular in the aquarium market due to its attractive coloured pattern of the body and special feeding behavior on starfish. For this reason, observations and records have been made on the reproductive habits, reproductive cycle and embryonic development of harlequin shrimp. The results showed that the mating behavior of the broodstock was dominated by males in four main stages of contact, climb and straddle, spermatophore transfer and separation. Egg spawning occurred 2 hours after mating. Under the temperature of (27 ± 0.5) ℃, embryos developed in 14 to 15 days, going through 8 stages of fertilized egg, cleavage, blastula, gastrula, pre-nauplius, post-nauplius, membrane-zoea and pre-hatching until larvae hatched. Females molted within 12 to 24 hours after larvae hatching and began mating again within 2 hours, and then entered the next breeding cycle. Our results provide not only a better understanding of the reproductive biology in Hymenocera picta, but also a useful reference for its artificial breeding practices.
Key words: Hymenocera picta; reproductive habits; embryonic development
ZHANG Shuyi , LIU Gaige , CHEN Jinmin , CHEN Nan , WANG Shuhong . Reproductive habit and embryonic development of the Harlequin Shrimp, Hymenocera picta (Decapoda: Hymenoceridae)[J]. Journal of Tropical Oceanography, 2025 , 44(2) : 92 -99 . DOI: 10.11978/2024088
表1 4对油彩蜡膜虾(Hymenocera picta)产卵情况统计(n=3)Tab. 1 Statistics of spawning in 4 pairs of Hymenocera picta (n=3) |
| 亲虾组别 | 产卵间隔/d | 产卵量/粒 |
|---|---|---|
| A | 16.33±0.58 | 992±118 |
| B | 16.67±0.58 | 637±93 |
| C | 17.00±1.00 | 1389±154 |
| D | 17.00±0.00 | 1192±186 |
| 平均值 | 16.75±0.62 | 1052±314 |
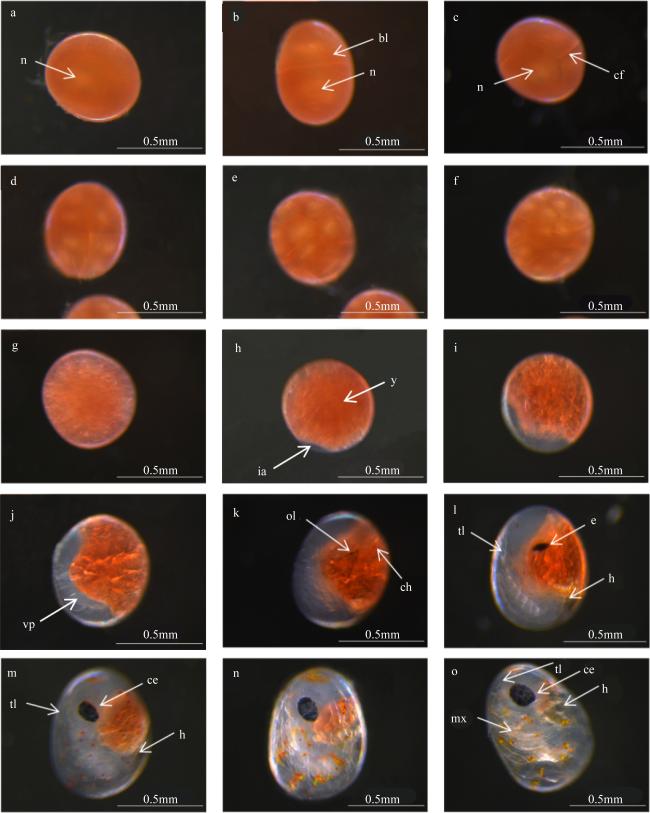
表2 油彩蜡膜虾(Hymenocera picta)胚胎发育时序Tab. 2 Time sequence of embryonic development in Hymenocera picta |
| 发育时期 | 持续时间 | 胚胎大小(长轴×短轴)/mm | 卵团颜色 | 主要特征 |
|---|---|---|---|---|
| 受精卵 | 2h | (0.56±0.21) × (0.44±0.34) | 褐色 | 充满卵黄, 不透明, 可以看见细胞核 |
| 卵裂期 | 20h | (0.57±0.14) × (0.45±0.17) | 褐色 | 出现卵裂沟, 细胞开始分裂 |
| 囊胚期 | 1d | (0.60±0.21) × (0.51±0.19) | 黄褐色 | 形成边围囊胚, 无囊胚腔, 也无法辨别动物极与植物极 |
| 原肠胚期 | 4d | (0.62±0.21) × (0.51±0.18) | 黄褐色 | 出现透明区域并逐渐扩大 |
| 前无节幼体期 | 3.5d | (0.68±0.16) × (0.55±0.11) | 浅褐色 | 透明区域出现分节 |
| 后无节幼体期 | 2d | (0.78±0.32) × (0.58±0.21) | 浅褐色 | 视叶出现, 尾节形成心脏形成, 存在心跳间隔 |
| 膜内溞状幼体期 | 2d | (0.86±0.26) × (0.62±0.12) | 浅褐色 | 视叶逐渐发育为复眼, 心脏跳动频率逐渐规律 |
| 孵化前期 | 1d | (0.91±0.29) × (0.64±0.14) | 浅灰色 | 卵黄被大量消耗, 复眼结构清晰, 附肢出现颤动 |
图 2 油彩蜡膜虾(Hymenocera picta)的胚胎发育过程a. 受精卵; b. 二细胞卵裂期; c. 四细胞卵裂期; d. 八细胞卵裂期; e. 十六细胞卵裂期; f. 多细胞卵裂期; g. 囊胚期; h~i. 原肠期; j. 前无节幼体期; k~l. 后无节幼体期; m~n. 膜内溞状幼体期; o. 孵化前期。bl: 卵裂球; ce: 复眼; cf: 卵裂沟; ch: 色素细胞; e: 眼; h: 心脏; ia: 内陷区; mx: 颚足; n: 细胞核; ol: 视叶; tl: 尾节; vp: 腹板; y: 卵黄 Fig. 2 Embryonic development stages of Hymenocera picta. (a) fertilized egg; (b to f) cleavage (b. 2-cell; c. 4-cell; d. 8-cell; e. 16-cell; f. multicell); (g) blastula; (h to i) gastrula; (j) pre-nauplius; (k to l) post-nauplius; (m to n) membrane-zoea stage; (o) pre-hatching. bl. blastomere; ce. compound eye; cf. cleavage furrow; ch. chromatophore; e. eye; h. heart; ia. invaginate area; mx. maxilliped; n. nucleolus; ol. ocular lobe; tl. telson; vp. ventral plate; y. yolk |
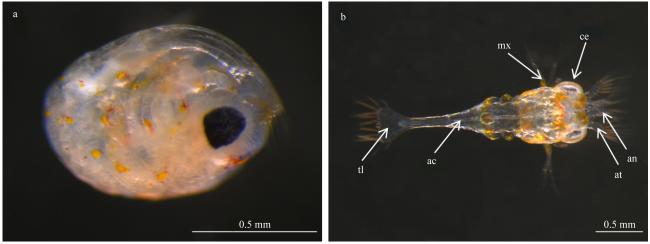
图 3 油彩蜡膜虾(Hymenocera picta)的孵化过程a. 正在破膜的胚胎; b. Ⅰ期溞状幼体。ac: 消化道; an: 第一触角; at: 第二触角; ce: 复眼; mx: 颚足; tl: 尾节 Fig. 3 Hatching development of Hymenocera picta. (a) Embryo during membrane rupture; (b) zoeal in Stage Ⅰ. ac: alimentary canal; an: antennule; at: antenna; ce: compound eye; mx: maxilliped; tl: telson |
| [1] |
邓杰文, 陈婉怡, 2022. 观赏中华锯齿米虾的培育与养殖研究进展[J]. 乡村科技, 13(18): 80-83 (in Chinese).
|
| [2] |
张雅静, 蔡生力, 刘红, 2012. 海洋观赏虾类清洁虾胚胎发育的形态学观察[J]. 中国水产科学, 19(6): 923-929.
|
| [3] |
张志峰, 廖承义, 王海林, 等, 1997. 中国对虾胚胎发育的研究[J]. 水产学报, 21(2): 201-205.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}