
Journal of Tropical Oceanography >
Observation of the fertilization process of Sargassum hemiphyllum var. chinense J. Agardhon Naozhou Island and a preliminary study on the optimal growth conditions for the juvenile sporophyte
Copy editor: YIN Bo
Received date: 2024-06-04
Revised date: 2024-07-02
Online published: 2024-07-09
Supported by
National Key Research and Development Program of China(2022YFD2401303)
2023 Major Science and Technology Research and Launch Project of Ningbo City(2023Z118)
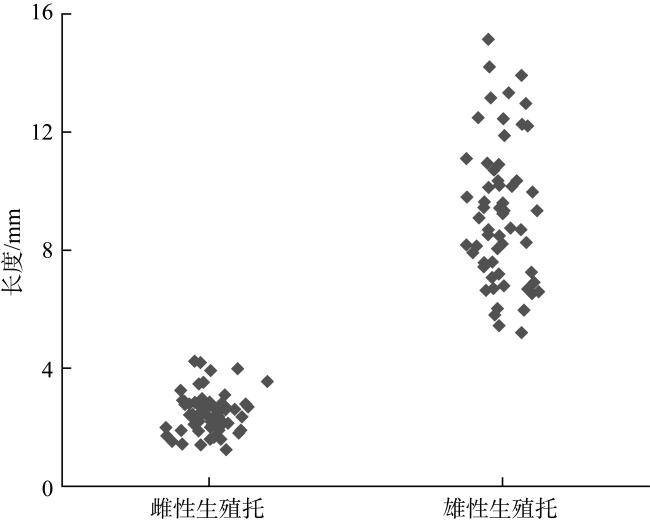
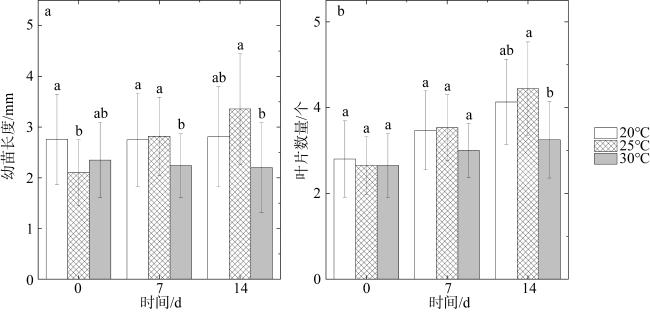
Sargassum hemiphyllum var. chinense J. Agardh is an economic seaweed widely distributed along the coast of the South China Sea. To investigate the sexual reproductive characteristics and the feasibility of artificial seedling propagation of this brown macroalga, this study employed optical microscopy to examine the morphological features of the thallus, male and female receptacles, and the early developmental stages of fertilized eggs in S. hemiphyllum var. chinense from Naozhou Island. The results revealed that the mature thallus of this Chinese variant exhibited a yellow-brown hue with a height of 0.8 to 1 m. It exhibited dioecious traits and possessed discoid holdfasts composed of repetitive bifurcations. The female receptacles were yellow-brown and cylindrical, adorned with numerous surface spines, measuring (0.46 ± 0.05) mm in diameter and (2.50 ± 0.67) mm in length. Conversely, the male receptacles were lighter in color, adopting a rod-like morphology, with a diameter of (1.01 ± 0.18) mm and a length of (9.21 ± 2.36) mm. Eggs released from the female structures exhibited an eight-nucleate one-egg configuration, either ellipsoidal or oval, with an outside diameter of (76.32 ± 10.11) μm. Following ovulation, these eggs adhered to the female structures awaiting fertilization. Sperms released from the male structures were pear-shaped, measuring (27.52 ± 5.43) μm in length and equipped with flagella. At the post-fertilization stage, the majority of fertilized eggs detached from the receptacles, affixing to substrates and undergoing early developmental processes. Within six hours, these fertilized eggs underwent their initial transverse division, subsequently progressing through multiple transverse and longitudinal divisions, and ultimately culminating in the development of landmine-shaped sporophytes within 24 hours, with primary fronds emerging after 28 days. Additionally, this study investigated the indoor use of suitable artificial substrates for the germination of sporophytes and the effect of temperature on the S. hemiphyllum var. chinense growth. Results indicated that, after seven days of culture, the mesh curtain yielded a germination rate of (7.25 ± 3.64) sporophytes per cm2, measuring (1.38 ± 0.35) mm in sporophytes’ length. Additional indoor suspension culture experiments revealed a 90% survival rate of sporophytes at the optimal temperature of 25℃, with a thallus growth rate of 90.1 μm·d−1, significantly outperforming other temperatures. These findings provide valuable preliminary data on which to base further studies of the early development and large-scale artificial seedling propagation of S. hemiphyllum var. chinense, thereby facilitating the rapid establishment of natural Sargassum habitats in coastal regions and contributing to the swift rehabilitation of China’s nearshore marine ecosystems.
YANG Wencheng , XIE Enyi , LIN Kun , GUO Youyou , WANG Huihui , CHEN Chunli , XIN Rong , CUI Jianjun . Observation of the fertilization process of Sargassum hemiphyllum var. chinense J. Agardhon Naozhou Island and a preliminary study on the optimal growth conditions for the juvenile sporophyte[J]. Journal of Tropical Oceanography, 2025 , 44(2) : 137 -146 . DOI: 10.11978/2024119
图2 硇洲岛半叶马尾藻中国变种成熟生殖托外观a. 成熟藻体(上雌下雄); b. 雌雄生殖托(左雌右雄); c. 挂卵的成熟雌性生殖托; d. 排精的成熟雄性生殖托 Fig. 2 Morphological features of mature receptacles of S. hemiphyllum var. chinense J. Agardh from Naozhou Island. The mature plants (above: female; below: male) (a); the receptacles (left: female; right: male) (b); eggs adhered to the female receptacle (c); sperms released by the male receptacle (d) |
图3 硇洲岛半叶马尾藻中国变种雌、雄生殖托长度Fig. 3 The length of female and male receptacles of S. hemiphyllum var. chinense J. Agardh from Naozhou Island |
表1 硇洲岛半叶马尾藻中国变种第二分支上生殖托数量、长度和直径Tab. 1 The number, length and diameter of the receptacles from the second lateral branches of S. hemiphyllum var. chinense J. Agardh from Naozhou Island |
| 性别 | 第二分支生殖托总数/个 | 生殖托长/mm | 生殖托直径/mm |
|---|---|---|---|
| 雄性 | 74.17 ± 12.69 | 9.21 ± 2.36 | 1.01 ± 0.18 |
| 雌性 | 56 ± 4.08 | 2.50 ± 0.67 | 0.46 ± 0.05 |
图5 硇洲岛半叶马尾藻变种受精卵发育过程a. 受精卵; b. 第一次横分裂后的受精卵; c. 分裂中的受精卵; d. “地雷状”幼孢子体; e. 萌发出假根的幼孢子体; f. 幼孢子体幼苗 Fig. 5 The early development of zygotes of S. hemiphyllum var. chinense J. Agardh from Naozhou Island. (a) Zygotes; (b) first division of zygotes across the longer axis; (c) zygotes after several divisions; (d) the landmine-shaped juvenile sporophytes formed after multiple divisions; (e) the sporophytes that geminated rhizoids; (f) juvenile sporophyte germlings |
图6 硇洲岛半叶马尾藻变种幼苗成功萌发的附着基a. 牡蛎壳上附着的幼孢子体; b. 布帘上附着的幼孢子体; c. 一个月后贝壳上的幼苗; d. 一个月后布帘上的幼苗 Fig. 6 Attachment materials for successful germination of S. hemiphyllum var. chinense J. Agardh from Naozhou Island. (a) Juvenile sporophytes attached to the oyster shells; (b) juvenile sporophytes attached to the curtain; (c) the 30 days’ germlings on the oyster shell; (d) the 30 days’ germlings on the curtain |
表2 硇洲岛半叶马尾藻中国变种不同附着基幼孢子体附着和幼苗生长情况Tab. 2 The attachment of juvenile sporophytes and growth of germlings for different attachment materials in the S. hemiphyllum var. chinense J. Agardh from Naozhou Island |
| 基质 | 幼孢子体附着 | 幼苗长度/mm | 幼苗密度/(indiv.·cm-2) |
|---|---|---|---|
| 牡蛎壳 | 有 | 0.92 ± 0.34a | 3.20 ± 2.15a |
| 布帘 | 有 | 1.38 ± 0.35b | 7.25 ± 3.64b |
| 铝板 | 无 | — | — |
| 塑料板 | 无 | — | — |
| 水泥砖 | 有 | — | — |
注: —表示无幼苗存活; 不同小写字母表示组别之间差异显著(P < 0.05) |
表3 不同温度条件下硇洲岛半叶马尾藻中国变种幼苗培养第7天和第14天的存活率和生长速率Tab. 3 Survival rate and growth rate of Naozhou Island S. hemiphyllum var. chinense J. Agardh germlings on the 7th and 14th days of cultivation under different temperature conditions. |
| 第7天 | 第14天 | |||||
|---|---|---|---|---|---|---|
| 20℃ | 25℃ | 30℃ | 20℃ | 25℃ | 30℃ | |
| 存活率/% | 75 | 95 | 55 | 75 | 90 | 40 |
| 生长速率/(μm·d-1) | -1.4 | 102.9 | -15.7 | 3.6 | 90.1 | -10.6 |
图7 不同温度对硇洲岛半叶马尾藻变种幼苗长度(a)和叶片数量(b)的影响不同小写字母表示组别之间的差异显著(P < 0.05) Fig. 7 Effects of different temperatures on the germlings’ length (a) and fronds’ number (b) of S. hemiphyllum var. chinense J. Agardh from Naozhou Island Different lowercase letters indicate significant differences between groups (P<0.05) |
| [1] |
韩军军, 白凯强, 赵明林, 等, 2016. 瓦氏马尾藻有性繁殖和幼孢子体发育的形态学观察[J]. 水生生物学报, 40(6): 1215-1220.
|
| [2] |
贺亮, 2017. 高温胁迫下半叶马尾藻中国变种生理生化响应及其耐热机制的初步研究[D]. 湛江: 广东海洋大学.
|
| [3] |
黄冰心, 丁兰平, 谭华强, 等, 2013. 我国沿海马尾藻属(Sargassum)的物种多样性及其区系分布特征[J]. 海洋与湖沼, 44(1): 69-76.
|
| [4] |
贾柽, 2012. 基于人工育苗的硇洲马尾藻增殖与移殖[D]. 湛江: 广东海洋大学.
|
| [5] |
贾柽, 杨彬, 谢恩义, 2012. 莫氏马尾藻繁殖生物学初步研究[J]. 水产科学, 31(10): 616-619.
|
| [6] |
姜森颢, 任贻超, 唐伯平, 等, 2017. 我国刺参养殖产业发展现状与对策研究[J]. 中国农业科技导报, 19(9): 15-23.
|
| [7] |
李爵乾, 廖嘉欣, 谢恩义, 2018. 温度对半叶马尾藻生长及生化组分的影响[J]. 河南农业, (5): 56, 59 (in Chinese).
|
| [8] |
林立东, 尚天歌, 张甜甜, 等, 2020. 褐藻羊栖菜繁殖生物学特征和生活史流程的补充研究[J]. 水产学报, 44(4): 581-595.
|
| [9] |
刘玮, 2015. 鼠尾藻群体资源调查及人工繁育的研究[D]. 北京: 中国科学院研究生院(海洋研究所).
|
| [10] |
骆其君, 2002. 赤潮对羊栖菜产量构成的影响[J]. 中国水产科学, 9(1): 95-96.
|
| [11] |
逄少军, 费修绠, 肖天, 等, 2000. 羊栖菜生殖托的离体培养的初步研究[J]. 海洋科学, 24(3): 1-3.
|
| [12] |
曲元凯, 谢恩义, 孙立伟, 等, 2015. 3种马尾藻周年生长和成熟时间以及形态学研究[J]. 安徽农业科学, 43(23): 136-139, 231.
|
| [13] |
孙建璋, 庄定根, 陈万东, 等, 2008. 铜藻Sargassum horneri繁殖生物学及种苗培育研究[J]. 南方水产, 4(2): 6-14.
|
| [14] |
孙立伟, 谢恩义, 曲元凯, 2015. 亨氏马尾藻的有性生殖观察[J]. 南方水产科学, 11(1): 62-67.
|
| [15] |
孙修涛, 王飞久, 汪文俊, 等, 2010. 基于有性繁殖的鼠尾藻规模化繁育试验[J]. 渔业科学进展, 31(3): 84-91.
|
| [16] |
田淑娴, 陈万东, 林利, 等, 2014. 繁殖期半叶马尾藻中国变种的形态结构观察[J]. 海洋渔业, 36(2): 107-115.
|
| [17] |
王惠, 谢恩义, 徐日升, 等, 2019. 硇洲马尾藻Sargassum naozhouense幼孢子体附着及原生态恢复[J]. 广东海洋大学学报, 39(1): 42-48.
|
| [18] |
向玉哲, 2022. 人工藻礁的选型与藻类附着效果的初步研究[D]. 汕头: 汕头大学.
|
| [19] |
邢珊珊, 邹潇潇, 林勇, 等, 2016. 匍枝马尾藻有性繁殖和人工育苗的初步研究[J]. 水产科学, 35(4): 376-380.
|
| [20] |
于沛民, 2007. 人工藻礁的选型与藻类附着效果的初步研究[D]. 青岛: 中国海洋大学.
|
| [21] |
曾呈奎, 2000. 中国海藻志. 第三卷褐藻门, 第二册墨角藻目[M]. 北京: 科学出版社.
|
| [22] |
曾昭钧, 孙立伟, 谢恩义, 2024. 围氏马尾藻周年生长及有性生殖的早期发育研究[J]. 热带海洋学报, 44(1): 44-52.
|
| [23] |
张立宁, 2013. 羊栖菜孢子体繁育及幼苗北方海区渡夏培育的研究[D]. 宁波: 宁波大学.
|
| [24] |
赵素芬, 2012. 海藻与海藻栽培学[M]. 北京: 国防工业出版社.
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}