
Journal of Tropical Oceanography >
Taxonomic research of the genus Hansarsia from the South China Sea*
Copy editor: YIN Bo
Received date: 2024-05-27
Revised date: 2024-08-09
Online published: 2024-08-14
Supported by
Capacity Building for Strategic Biological Resources, the Chinese Academy of Sciences(KFJ-BRP-017-48)
Taxonomist project, Chinese Academy of Sciences(CAS-TAX-24-042)
National Animal Collection Resource Center, China
Guangdong Provincial Observation and Research Station for Tropical Ocean Environment in Western Coastal Waters(GSTOEW)
National Natural Science Foundation of China(42225602)
National Natural Science Foundation of China(92258301)
National Natural Science Foundation of China(42090042)
Species of the genus Hansarsia play crucial roles in marine planktonic ecosystems, but there is often confusion and misidentification. This paper employed morphological analysis methods to conduct in-depth research on the specimens of the genus Hansarsia collected by the Marine Biodiversity Collections of the South China Sea, Chinese Academy of Sciences, from the South China Sea and the East Indian Ocean, aiming to clarify differences between species and resolve taxonomic confusions. This paper a) supplemented and revised the characterizations of H. tenella, H. gracilis, H. microps, and H. atlantica; b) depicted the changes in the first and second thoracic legs of the four species from juvenile to adult; c) pointed out the phenomenon of two rostrum shapes in the adult H. gracilis, and the variation in the number of setae on the propodus of the first thoracic leg; and d) clarified the distinctions between H. microps and H. atlantica in the South China Sea and the East Indian Ocean.
Key words: krill; Hansarsia; morphology; taxonomy; the first thoracic leg
ZHOU Zhiqian , LIAN Xiping , TAN Yehui . Taxonomic research of the genus Hansarsia from the South China Sea*[J]. Journal of Tropical Oceanography, 2025 , 44(2) : 100 -114 . DOI: 10.11978/2024110
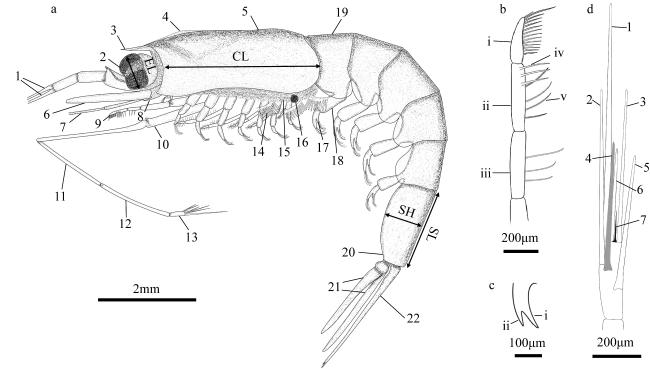
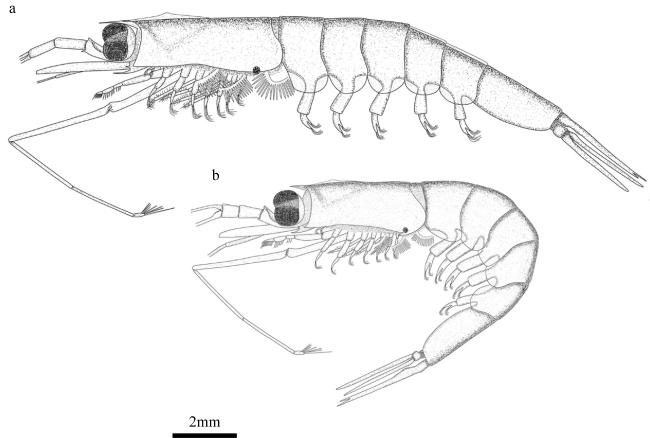
图1 常用术语明细a. 瘦细足磷虾, 长型额角雌性(12mm, 南海)。1. 第一触角柄; 2. 第一触角鞭; 3. 额角; 4. 头胸甲棱; 5. 头胸甲; 6. 鳞片; 7. 第二触角柄; 8. 第二触角柄部腹侧刺; 9. 第一胸足; 10. 第二胸足; 11. 腕节; 12. 掌节; 13. 指节; 14. 指状足鳃; 15. 雌性交接器/体外纳精器; 16. 发光器; 17. 雄性交接器; 18. 第一腹足; 19. 第一腹节; 20. 肛前刺; 21. 尾肢; 22. 尾节。EL: 眼高; CL: 头胸甲长度; SH: 第六腹节宽度; SL: 第六腹节长度。b. 第一胸足末部。i. 指节; ii. 掌节; iii. 腕节; iv. 掌节内排刚毛; v. 掌节外排刚毛。c. 肛前刺。i. 内叉; ii. 外叉。d. 第二胸足指节。图a和图d中数字表示刺毛生长先后顺序 Fig. 1 List of commonly used terms. (a) Hansarsia gracilis, female with long rostrum (12 mm, the South China Sea). 1. antennular peduncle; 2. antennular flagella; 3. rostrum; 4. carapace keel; 5. carapace; 6. scale; 7. antennal peduncle; 8. ventrolateral spine on coxa of antenna; 9. the first thoracic leg; 10. the second thoracic leg; 11. carpus; 12. propodus; 13. dactylus; 14. podobranchia; 15. thelyca (female); 16. photophore; 17. petasma (male); 18. the first pleopod; 19. the first abdominal segment; 20. pre-anal spine; 21. uropod; 22. telson; EL: eye length; CL: carapace length; SH: height of the sixth pleonic somite; SL: length of the sixth pleonic somite. (b) distal part of first thoracic leg. i. dactylus; ii. propodus; iii. carpus; iv. the inner setae of propodus; v. the exteral setae of propodus. (c) pre-anal spine. i. inner fork; ii. external fork. (d) dactylus of second thoracic leg. numbers in panels (a) and (d) indicate the order of setae growth |
表1 秀细足磷虾标本测量Tab. 1 Specimen measurement of Hansarsia tenella |
| 采样海域 | 发育状态 | 性别 | 体长/mm | 眼/mm | 额角/mm | 头胸甲/mm |
|---|---|---|---|---|---|---|
| 东印度洋 | 成体 | 雌性 | 20.464 | 1.334 | 1.181 | 4.988 |
| 东印度洋 | 成体 | 雌性 | 16.657 | 1.017 | 0.804 | 4.148 |
| 东印度洋 | 成体 | 雌性 | 21.755 | 1.423 | 1.234 | 5.303 |
| 南海 | 亚成体 | 雌性 | 9.744 | 0.652 | 0.432 | 2.140 |
| 南海 | 成体 | 雌性 | 12.678 | 0.849 | 0.971 | 2.930 |
| 东印度洋 | 亚成体 | 雌性 | 10.398 | 0.720 | 0.948 | 2.151 |
| 南海 | 亚成体 | 雌性 | 8.458 | 0.534 | 0.505 | 2.158 |
| 南海 | 亚成体 | 雌性 | 9.537 | 0.644 | 0.686 | 2.235 |
| 东印度洋 | 成体 | 雌性 | 12.765 | 0.867 | 0.670 | 3.105 |
| 南海 | 亚成体 | 雌性 | 10.406 | 0.675 | 0.601 | 2.516 |
| 南海 | 成体 | 雌性 | 12.961 | 0.857 | 0.720 | 3.132 |
| 南海 | 亚成体 | 雌性 | 9.765 | 0.644 | 0.529 | 2.540 |
| 东印度洋 | 成体 | 雄性 | 16.287 | 0.922 | 0.800 | 3.664 |
| 东印度洋 | 成体 | 雄性 | 18.123 | 1.027 | 0.870 | 4.122 |
| 南海 | 成体 | 雄性 | 16.116 | 0.994 | 0.900 | 3.370 |
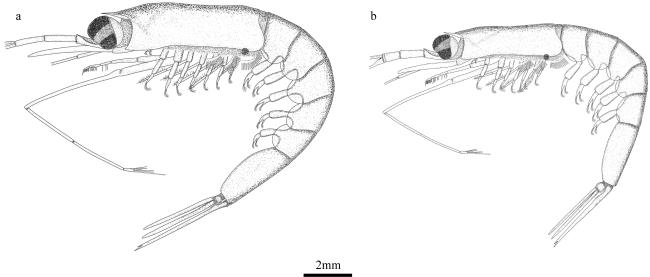
图4 雄性交接器a. 秀细足磷虾。1. 辅叶; 2. 中叶; 3. 内叶; 4. 侧突; 5. 基突; 6. 顶突; 7. 刺状突。b. 瘦细足磷虾。i. 南海常见, ii和iii. 东印度洋常见。c. 小细足磷虾。d. 长细足磷虾 Fig. 4 Petasma. (a) Hansarsia tenella. 1. auxillary lobe; 2. median lobe; 3. inner lobe; 4. lateral process; 5. proximal process; 6. terminal process; 7. spine-shaped process. (b) Hansarsia gracilis. i. Commonly found in the South China Sea, ii. and iii. commonly found in the East Indian Ocean; (c) Hansarsia microps; (d) Hansarsia atlantica |
表2 瘦细足磷虾标本测量Tab. 2 Specimen measurement of Hansarsia gracilis |
| 采样海域 | 发育状态 | 性别 | 体长/mm | 眼/mm | 额角/mm | 头胸甲/mm |
|---|---|---|---|---|---|---|
| 东印度洋 | 成体 | 雌性 | 10.075 | 0.607 | 0.788 | 3.210 |
| 东印度洋 | 亚成体 | 雌性 | 9.045 | 0.663 | 0.775 | 2.111 |
| 东印度洋 | 亚成体 | 雌性 | 8.786 | 0.511 | 0.673 | 2.151 |
| 南海 | 亚成体 | 雌性 | 9.210 | 0.651 | 0.544 | 2.376 |
| 东印度洋 | 亚成体 | 雌性 | 9.445 | 0.658 | 0.762 | 2.422 |
| 东印度洋 | 成体 | 雌性 | 15.248 | 0.968 | 0.899 | 4.326 |
| 南海 | 成体 | 雌性 | 11.985 | 0.669 | 0.665 | 2.799 |
| 南海 | 亚成体 | 雌性 | 9.732 | 0.638 | 0.548 | 2.542 |
| 南海 | 成体 | 雌性 | 12.350 | 0.844 | 0.768 | 3.294 |
| 东印度洋 | 成体 | 雌性 | 12.338 | 0.777 | 0.692 | 3.083 |
| 南海 | 亚成体 | 雌性 | 10.651 | 0.684 | 0.671 | 2.412 |
| 南海 | 成体 | 雌性 | 13.010 | 0.795 | 0.787 | 3.470 |
| 东印度洋 | 成体 | 雌性 | 14.154 | 0.822 | 0.748 | 3.781 |
| 东印度洋 | 成体 | 雌性 | 9.941 | 0.689 | 0.707 | 2.689 |
| 南海 | 成体 | 雌性 | 11.077 | 0.547 | 0.582 | 2.822 |
| 南海 | 成体 | 雌性 | 11.916 | 0.577 | 0.820 | 2.969 |
| 南海 | 成体 | 雌性 | 10.887 | 0.575 | 0.644 | 2.545 |
| 东印度洋 | 成体 | 雌性 | 13.268 | 0.843 | 0.841 | 3.507 |
| 南海 | 成体 | 雌性 | 13.654 | 0.816 | 0.758 | 3.760 |
| 南海 | 成体 | 雌性 | 13.262 | 0.892 | 0.693 | 3.806 |
| 南海 | 成体 | 雌性 | 9.732 | 0.717 | 0.555 | 2.390 |
| 南海 | 成体 | 雌性 | 13.095 | 0.898 | 0.582 | 3.448 |
| 南海 | 成体 | 雌性 | 11.490 | 0.822 | 0.462 | 2.994 |
| 南海 | 成体 | 雌性 | 13.879 | 0.927 | 0.597 | 3.694 |
| 东印度洋 | 成体 | 雌性 | 12.281 | 0.873 | 0.538 | 3.248 |
| 南海 | 成体 | 雌性 | 11.552 | 0.788 | 0.463 | 3.033 |
| 东印度洋 | 成体 | 雌性 | 13.819 | 0.937 | 0.609 | 3.665 |
| 东印度洋 | 成体 | 雌性 | 13.431 | 0.883 | 0.462 | 3.478 |
| 东印度洋 | 成体 | 雌性 | 14.655 | 0.920 | 0.550 | 4.245 |
| 东印度洋 | 成体 | 雌性 | 14.530 | 0.927 | 0.584 | 4.083 |
| 东印度洋 | 成体 | 雌性 | 14.558 | 0.923 | 0.559 | 4.238 |
| 东印度洋 | 成体 | 雌性 | 14.758 | 0.710 | 0.481 | 4.194 |
| 南海 | 成体 | 雌性 | 13.260 | 0.863 | 0.497 | 3.818 |
| 南海 | 成体 | 雌性 | 13.029 | 0.931 | 0.582 | 3.767 |
| 东印度洋 | 成体 | 雌性 | 14.074 | 0.925 | 0.624 | 4.021 |
| 东印度洋 | 成体 | 雌性 | 13.682 | 0.859 | 0.581 | 4.110 |
| 东印度洋 | 成体 | 雌性 | 13.720 | 0.834 | 0.552 | 3.649 |
| 东印度洋 | 成体 | 雌性 | 13.978 | 0.920 | 0.469 | 3.966 |
| 东印度洋 | 成体 | 雌性 | 13.905 | 0.907 | 0.601 | 4.167 |
| 南海 | 成体 | 雌性 | 12.748 | 0.984 | 0.543 | 3.437 |
| 东印度洋 | 亚成体 | 雄性 | 9.235 | 0.618 | 0.622 | 2.295 |
| 南海 | 成体 | 雄性 | 10.003 | 0.715 | 0.590 | 2.514 |
| 南海 | 成体 | 雄性 | 10.880 | 0.725 | 0.558 | 2.874 |
| 南海 | 成体 | 雄性 | 10.316 | 0.698 | 0.552 | 2.400 |
| 南海 | 成体 | 雄性 | 10.781 | 0.747 | 0.561 | 2.550 |
| 南海 | 成体 | 雄性 | 10.874 | 0.656 | 0.766 | 2.586 |
| 南海 | 成体 | 雄性 | 11.607 | 0.763 | 0.517 | 2.856 |
| 南海 | 成体 | 雄性 | 10.913 | 0.715 | 0.624 | 2.662 |
| 南海 | 成体 | 雄性 | 10.645 | 0.749 | 0.501 | 2.665 |
| 南海 | 成体 | 雄性 | 11.258 | 0.650 | 0.309 | 2.565 |
| 东印度洋 | 成体 | 雄性 | 11.781 | 0.780 | 0.374 | 2.978 |
| 南海 | 成体 | 雄性 | 11.030 | 0.799 | 0.390 | 2.827 |
| 南海 | 成体 | 雄性 | 11.606 | 0.790 | 0.406 | 2.960 |
| 南海 | 成体 | 雄性 | 11.623 | 0.816 | 0.328 | 3.113 |
| 东印度洋 | 成体 | 雄性 | 11.904 | 0.769 | 0.303 | 2.983 |
| 南海 | 成体 | 雄性 | 10.894 | 0.767 | 0.400 | 2.840 |
| 南海 | 成体 | 雄性 | 11.663 | 0.835 | 0.334 | 2.908 |
| 东印度洋 | 成体 | 雄性 | 10.925 | 0.778 | 0.406 | 2.675 |
| 南海 | 成体 | 雄性 | 12.089 | 0.768 | 0.338 | 2.895 |
| 东印度洋 | 成体 | 雄性 | 11.904 | 0.749 | 0.412 | 3.051 |
| 东印度洋 | 亚成体 | 雄性 | 9.606 | 0.716 | 0.550 | 2.428 |
| 东印度洋 | 亚成体 | 雄性 | 9.816 | 0.731 | 0.604 | 2.561 |
| 东印度洋 | 成体 | 雄性 | 10.277 | 0.713 | 0.676 | 2.409 |
| 南海 | 成体 | 雄性 | 10.500 | 0.633 | 0.657 | 2.392 |
| 南海 | 成体 | 雄性 | 9.959 | 0.620 | 0.593 | 2.293 |
| 南海 | 成体 | 雄性 | 11.038 | 0.685 | 0.499 | 2.598 |
| 南海 | 亚成体 | 雄性 | 9.370 | 0.701 | 0.408 | 2.287 |
表3 小细足磷虾标本测量Tab. 3 Specimen measurement of Hansarsia microps |
| 采样海域 | 发育状态 | 性别 | 体长/mm | 眼/mm | 额角/mm | 头胸甲/mm |
|---|---|---|---|---|---|---|
| 东印度洋 | 成体 | 雌性 | 14.340 | 1.087 | 1.251 | 2.989 |
| 南海 | 亚成体 | 雌性 | 7.620 | 0.610 | 0.838 | 1.626 |
| 南海 | 亚成体 | 雌性 | 9.415 | 0.614 | 0.836 | 1.877 |
| 东印度洋 | 亚成体 | 雌性 | 10.213 | 0.717 | 0.864 | 2.268 |
| 东印度洋 | 亚成体 | 雌性 | 9.621 | 0.667 | 0.989 | 1.850 |
| 东印度洋 | 亚成体 | 雌性 | 8.792 | 0.658 | 0.848 | 1.935 |
| 南海 | 亚成体 | 雌性 | 8.323 | 0.631 | 0.734 | 1.696 |
| 南海 | 亚成体 | 雌性 | 8.229 | 0.727 | 0.905 | 1.903 |
| 南海 | 亚成体 | 雌性 | 7.682 | 0.628 | 0.951 | 1.491 |
| 东印度洋 | 亚成体 | 雌性 | 7.727 | 0.631 | 0.842 | 1.652 |
| 南海 | 成体 | 雌性 | 11.437 | 0.930 | 1.109 | 2.431 |
| 南海 | 成体 | 雌性 | 15.504 | 0.985 | 1.321 | 3.510 |
| 东印度洋 | 成体 | 雌性 | 10.430 | 0.776 | 0.880 | 2.165 |
| 东印度洋 | 成体 | 雌性 | 12.274 | 0.865 | 1.033 | 2.638 |
| 东印度洋 | 成体 | 雌性 | 17.876 | 1.036 | 1.248 | 4.407 |
| 东印度洋 | 成体 | 雌性 | 12.931 | 0.865 | 1.160 | 2.967 |
| 东印度洋 | 成体 | 雌性 | 14.152 | 0.868 | 1.163 | 2.934 |
| 东印度洋 | 亚成体 | 雌性 | 10.488 | 0.736 | 0.978 | 2.103 |
| 东印度洋 | 成体 | 雌性 | 17.899 | 1.118 | 1.373 | 4.206 |
| 东印度洋 | 成体 | 雌性 | 16.500 | 1.158 | 1.157 | 3.932 |
| 东印度洋 | 成体 | 雌性 | 11.677 | 0.883 | 0.974 | 2.574 |
| 东印度洋 | 成体 | 雌性 | 13.788 | 0.934 | 1.017 | 2.841 |
| 东印度洋 | 成体 | 雄性 | 14.765 | 0.847 | 1.269 | 3.087 |
| 东印度洋 | 成体 | 雄性 | 15.437 | 1.113 | 0.761 | 3.504 |
表4 长细足磷虾标本测量Tab. 4 Specimen measurement of Hansarsia atlantica |
| 采样海域 | 发育状态 | 性别 | 体长/mm | 眼/mm | 额角/mm | 头胸甲/mm |
|---|---|---|---|---|---|---|
| 东印度洋 | 成体 | 雌性 | 11.113 | 0.583 | 0.645 | 2.889 |
| 南海 | 成体 | 雌性 | 9.128 | 0.619 | 0.676 | 2.144 |
| 南海 | 成体 | 雌性 | 9.112 | 0.549 | 0.836 | 2.196 |
| 南海 | 成体 | 雌性 | 9.923 | 0.676 | 0.909 | 2.456 |
| 南海 | 成体 | 雌性 | 9.260 | 0.632 | 0.702 | 2.120 |
| 南海 | 亚成体 | 雌性 | 8.821 | 0.488 | 0.788 | 2.019 |
| 东印度洋 | 亚成体 | 雌性 | 7.203 | 0.463 | 0.652 | 1.731 |
| 南海 | 亚成体 | 雌性 | 6.637 | 0.409 | 0.535 | 1.588 |
| 东印度洋 | 亚成体 | 雌性 | 7.905 | 0.586 | 0.725 | 1.870 |
| 南海 | 亚成体 | 雌性 | 8.398 | 0.475 | 0.773 | 1.836 |
| 东印度洋 | 成体 | 雌性 | 9.102 | 0.635 | 0.665 | 2.219 |
| 南海 | 亚成体 | 雌性 | 8.807 | 0.554 | 0.655 | 2.069 |
| 南海 | 亚成体 | 雌性 | 8.168 | 0.536 | 0.703 | 1.920 |
| 东印度洋 | 亚成体 | 雌性 | 8.232 | 0.464 | 0.733 | 2.026 |
| 东印度洋 | 亚成体 | 雌性 | 7.871 | 0.539 | 0.722 | 1.919 |
| 东印度洋 | 亚成体 | 雌性 | 8.935 | 0.531 | 0.618 | 2.356 |
| 南海 | 成体 | 雌性 | 10.305 | 0.664 | 0.769 | 2.754 |
| 南海 | 成体 | 雄性 | 9.647 | 0.618 | 0.712 | 2.382 |
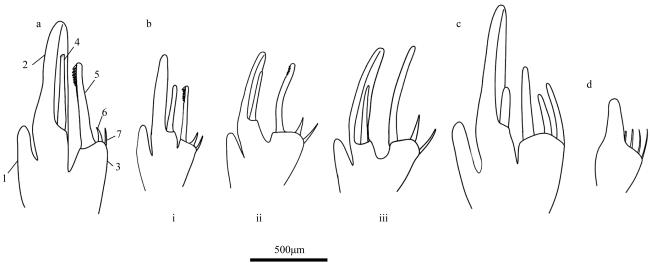
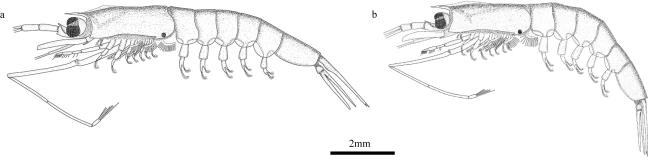
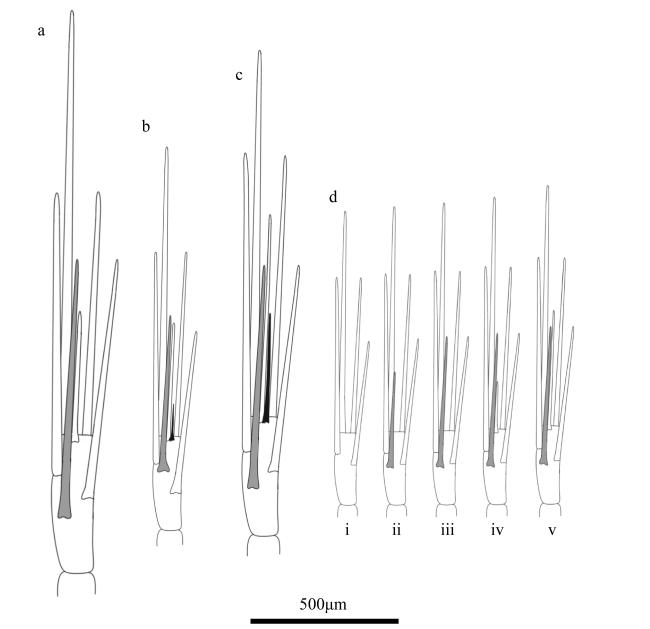
图8 第一胸足末部形态a. 瘦细足磷虾; b. 长细足磷虾; c. 小细足磷虾; d. 秀细足磷虾。*为雄性样本 Fig. 8 Morphology of distal part of first thoracic leg. (a) Hansarsia gracilis; (b) Hansarsia atlantica; (c) Hansarsia microps; (d) Hansarsia tenella. * for male samples |
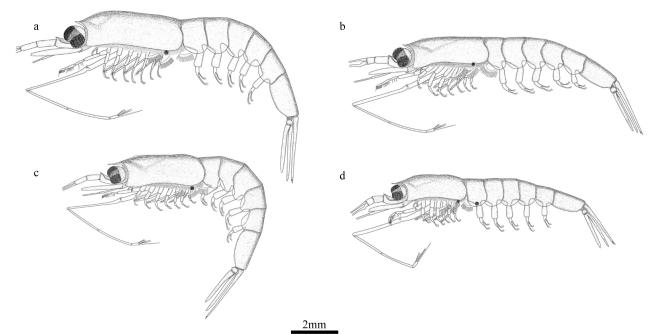
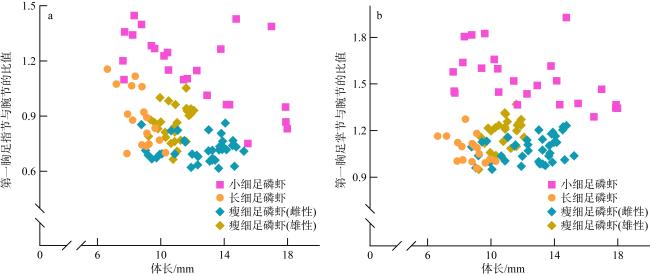
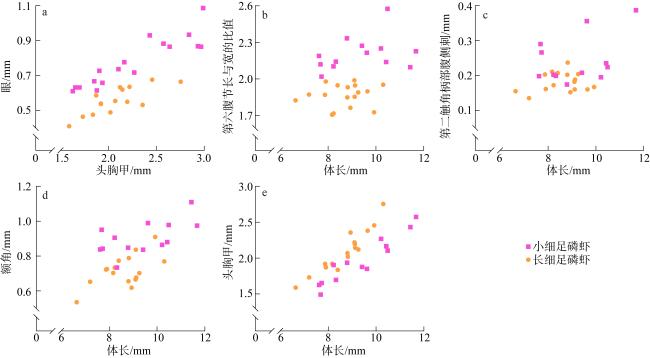
图10 小细足磷虾与长细足磷虾的形态特征a. 眼; b. 第六腹节长与宽的比值; c. 第二触角柄部腹侧刺; d. 额角; e. 头胸甲 Fig. 10 Morphological characteristics of Hansarsia microps and Hansarsia atlantica. (a) eye; (b) the proportions of length to width of the sixth abdominal segment; (c) rostrum; (d) ventrolateral spine on coxa of antenna; (e) carapace |
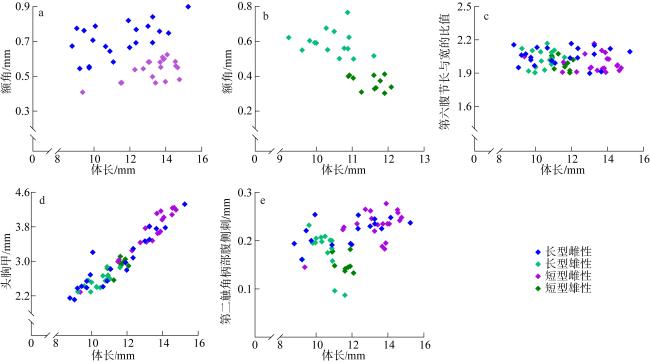
图11 瘦细足磷虾的形态特征a. 雄性额角; b. 雌性额角; c. 第六腹节长与宽的比值; d. 头胸甲; e. 第二触角柄部腹侧刺 Fig. 11 Morphological characteristics of Hansarsia gracilis. (a) male rostrum; (b) female rostrum; (c) the proportions of length to width of the sixth abdominal segment; (d) carapace; (e) ventrolateral spine on coxa of antenna |
| [1] |
蔡秉及, 1983. 太平洋热带水域磷虾类的初步研究[J]. 海洋通报, (2): 71-87 (in Chinese).
|
| [2] |
陈清潮, 张谷贤, 1991. 南海北部和中部的磷虾类[M]//中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物研究论文集(二). 北京: 科学出版社: 139-171 (in Chinese).
|
| [3] |
陈清潮, 2008. 磷虾目[M]//刘瑞玉. 中国海洋生物名录. 北京: 科学出版社: 699-701 (in Chinese).
|
| [4] |
李少菁, 陈清潮, 2012. 磷虾目[M]//黄宗国, 林茂. 中国海洋物种和图集上卷: 中国海洋物种多样性下册. 北京: 海洋出版社: 776-778 (in Chinese).
|
| [5] |
齐钟彦, 1999. 使用说明[M]//齐钟彦. 新拉汉无脊椎动物名称. 北京: 科学出版社:ix-x (in Chinese).
|
| [6] |
张谷贤, 陈清潮, 1991. 南海及其邻近海区的磷虾类[M]//中国科学院南沙综合科学考察队. 南沙群岛海区海洋动物区系和动物地理研究专集. 北京: 海洋出版社: 140-270 (in Chinese).
|
| [7] |
郑重, 李少菁, 许振祖, 1984. 磷虾类[M]//郑重, 李少菁, 许振祖. 海洋浮游生物学. 北京: 海洋出版社: 418-439 (in Chinese).
|
| [8] |
郑重, 李少菁, 郭东晖, 2011. 种类组成[M]//郑重, 李少菁, 郭东晖. 海洋磷虾类生物学. 厦门: 厦门大学出版社: 14-33 (in Chinese).
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}