
Journal of Tropical Oceanography >
Morphological classification and phagocytosis of haemocytes in Spondylus sp.
Copy editor: YIN Bo
Received date: 2024-05-25
Revised date: 2024-07-22
Online published: 2024-08-14
Supported by
Guangdong Provincial Department of Education Marine Shellfish Ecological Aquaculture and Disease Prevention and Control Innovation Team Project(2021KCXTD057)
Professors and doctors of Huizhou University started the project(15604220001)
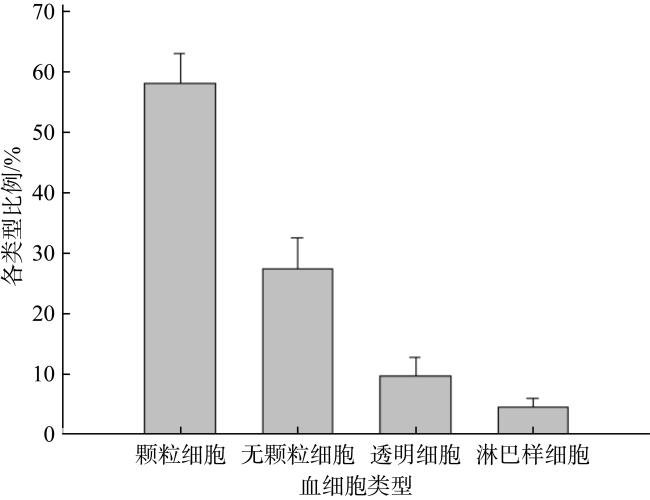
Blood cells, as the main immune effector cells, play a very important role in the immune defense system of shellfish. The morphology, type and phagocytosis function of hemocytes of Spondylus sp. were studied by means of blood cell count, Giemsa staining, transmission electron microscopy, scanning electron microscopy and fluorescence phagocytosis. The results showed that the blood cell density of the Spondylus sp. was 3.296 × 106 cells·mL-1, and the blood cells differ significantly in size and with regard to whether they contain particles within the cytoplasm. Based on these differences, they are subdivided them into four types: hyalinocyte (9.74%), granulocyte (58.14%), agranular haemocyte (27.50%) and lymphoid (4.62%). Hyalinocytes, agranular haemocytes, granulocytes and lymphoids could be observed under transmission electron microscope. Under the observation of scanning electron microscope, four different types of cells can be clearly distinguished, which are oval cells, round cells, spindle cells, nucleated (conical) cells. A phagocytosis experiment was carried out with Escherichia coli BL21 labeled with the green fluorescent protein (GFP), and the results showed that the phagocytosis of the blood cells from Spondylus sp. was mainly dependent on granulocytes.
ZHOU Kexin , ZHEN Wenjing , ZHANG Wenwen , WANG Jiangyong . Morphological classification and phagocytosis of haemocytes in Spondylus sp.[J]. Journal of Tropical Oceanography, 2025 , 44(2) : 84 -91 . DOI: 10.11978/2024108
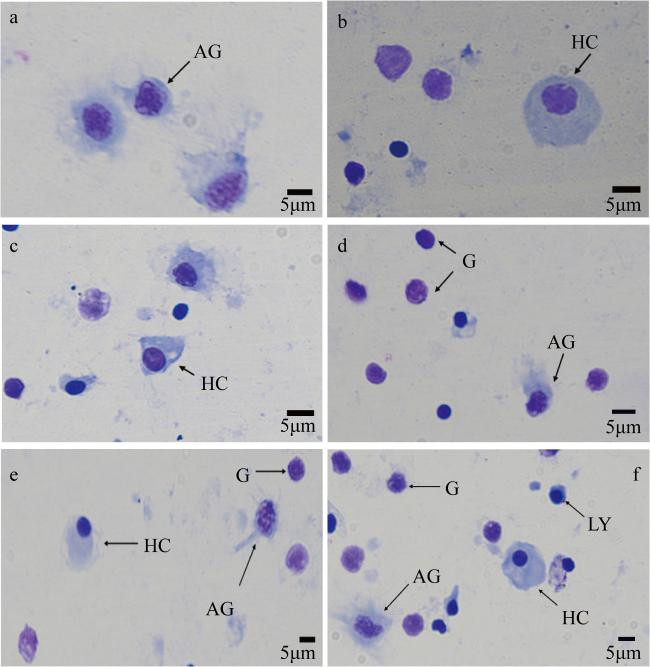
图1 海菊蛤血细胞光学显微镜观察(1000 ×)G: 颗粒细胞; AG: 无颗粒细胞; HC: 透明细胞; LY: 淋巴样细胞 Fig. 1 Optical microscope observation of haemocytes in Spondylus sp. (1000 ×). G: granulocyte; AG: agranular haemocyte; HC: hyalinocyte; LY: lymphoid haemocyte |
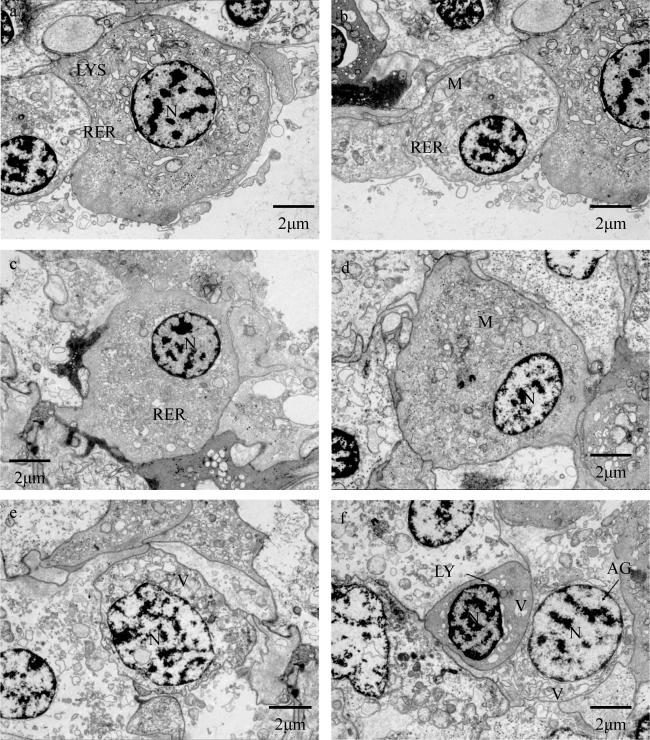
图3 海菊蛤血细胞透射电子显微镜观察(4000 ×)a, b. 颗粒细胞; c, d. 透明细胞; e, f. 淋巴样细胞; f. 无颗粒细胞。N: 细胞核; RER: 粗面内质网; LYS: 溶酶体; M: 线粒体; V: 液泡; LY: 淋巴样细胞; AG: 无颗粒细胞 Fig. 3 Morphology of haemocytes in Spondylus sp. by transmission electron microscope (4000 ×). a, b: granulocyte; c, d: hyalinocyte; e, f: lymphoid haemocyte; f: agranulocyte. N: nucleus; RER: rough endoplasmic reticulum; LYS: lysosome; M: mitochondria; V: vacuole; LY: lymphoid haemocyte; AG: agranular haemocyte |
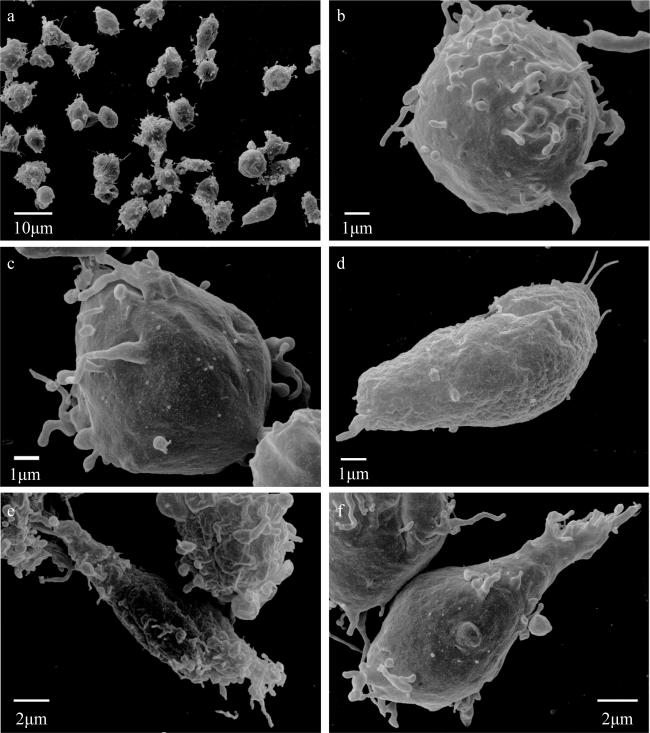
图4 海菊蛤血细胞的形态学-扫描电子显微镜观察a. 海菊蛤细胞(1500 ×); b. 圆形细胞(10000 ×); c. 椭圆细胞(10000 ×); d. 梭形细胞(10000 ×); e. 梭形细胞(7000 ×); f. 核形(锥形)细胞(8000 ×) Fig. 4 Morphology of haemocytes in Spondylus sp. by scanning electron microscope. a: spondylidae cells (1500 ×); b: round cells (10000 ×); c: elliptical cells (10000 ×); d: spindle cells (10000 ×); e: spindle cells (7000 ×); f: nucleated (conical) cells (8000 ×) |
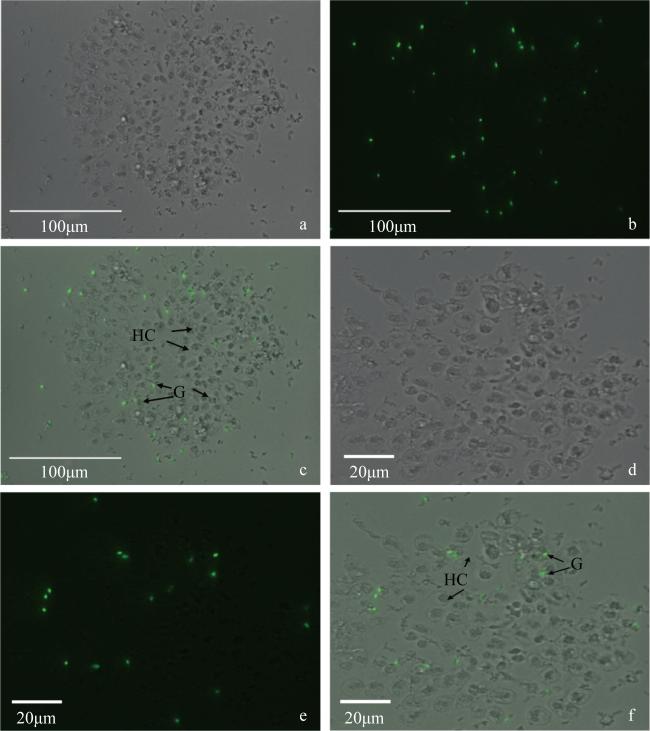
图5 海菊蛤血细胞的吞噬实验-荧光显微镜观察a, d. 明场/海菊蛤血细胞(400 ×); b, e. GFP/大肠杆菌BL21 (400 ×); c, f. Merge (400 ×)。G: 颗粒细胞; HC: 透明细胞 Fig. 5 Phagocytosis experiment and fluorescence microscope observation of haemocytes in Spondylus sp.. a, d: Bright field/Spondylus sp. haemocyte (400 ×); b, e: GFP/Escherichia coli BL21 (400 ×); c, f: Merge (400 ×). G: granulocyte; HC: hyalinocyte |
| [1] |
郭磊, 2007. 池蝶蚌血细胞类型及吞噬活性的研究[D]. 南昌: 南昌大学.
|
| [2] |
裴鹏祖, 胡宝庆, 谢彦海, 等, 2011. 中华圆田螺血细胞的分类和吞噬活性[J]. 南昌大学学报(理科版), 35(1): 83-89, 94.
|
| [3] |
宋小瑞, 2016. 长牡蛎免疫系统的发生和发育机制的初步研究[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
| [4] |
孙虎山, 李光友, 2003. 栉孔扇贝血细胞的分类[J]. 海洋学报, 25(S2): 51-57.
|
| [5] |
王江勇, 郭志勋, 冯娟, 等, 2010. 杂色鲍血细胞免疫特点及免疫功能的研究[J]. 热带海洋学报, 29(3): 71-76.
|
| [6] |
谢彦海, 2007. 褶纹冠蚌血细胞及其免疫特性研究[D]. 南昌: 南昌大学.
|
| [7] |
邢婧, 2002. 栉孔扇贝(Chlamys farreri)血细胞及其免疫特性的研究[D]. 青岛: 中国海洋大学.
|
| [8] |
杨军, 石安静, 2002. 3种淡水育珠河蚌血细胞类型的研究[J]. 四川大学学报(自然科学版), 39(S1): 68-72.
|
| [9] |
杨丽佳, 武梅, 王信超, 等, 2011. 厚壳贻贝血细胞及血清抗菌肽的分析[J]. 中国农学通报, 27(23): 58-64.
|
| [10] |
杨新春, 2006. 扇贝血细胞的形态学研究[D]. 青岛: 中国海洋大学.
|
| [11] |
朱泽闻, 徐立蒲, 吴信忠, 等, 2011. 泥蚶血细胞的形态结构特征及部分免疫功能[J]. 水产学报, 35(10): 1494-1504.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}