
Journal of Tropical Oceanography >
Study on biomass models of juvenile mangroves and carbon storage in young mangrove ecosystems*
Received date: 2024-07-18
Revised date: 2024-08-02
Online published: 2024-09-03
Supported by
Science and Technology Projects of Guangdong Provincial Department of Natural Resources(GDZRZYKJ2023002)
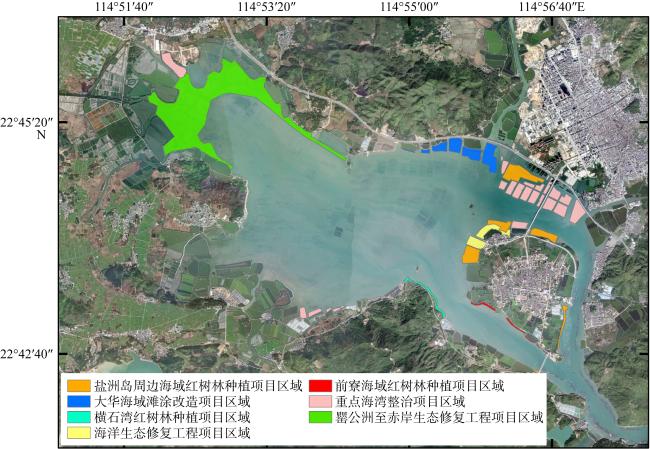
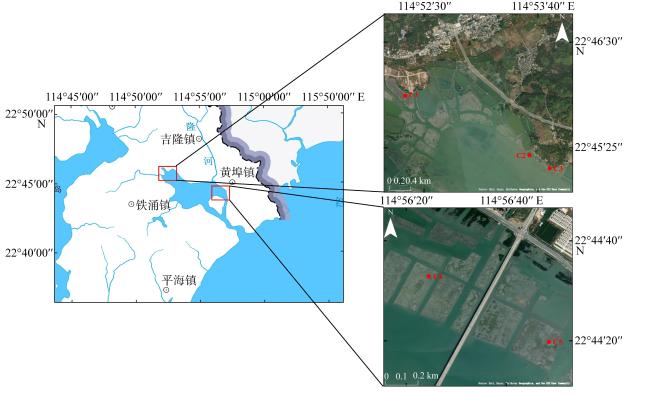
As global climate change intensifies, mangroves—a vital coastal blue carbon ecosystem—have garnered increasing attention. This study aimed to develop biomass models for juvenile mangroves and assess the carbon storage of young mangrove ecosystems in Kaozhouyang Bay. The results provide empirical methods and a scientific basis for the rapid and accurate assessment of carbon stocks in artificially planted young mangroves. The study focuses on five artificially planted juvenile mangrove species in Kaozhouyang Bay: Avicennia marina, Rhizophora stylosa, Kandelia obovata, Bruguiera gymnorhiza, and Aegiceras corniculatum. Various factors derived from basal diameter (D) and tree height (H) were used to construct optimal allometric equations between biomass and dendrometric parameters. Furthermore, the best-performing biomass models were applied to estimate vegetation carbon storage and ecosystem carbon storage in the artificially planted mangroves of Kaozhouyang Bay. The findings indicated that multivariable models generally outperformed univariable models, except for the below-ground biomass model of Aegiceras corniculatum. The optimal biomass models for Avicennia marina, Rhizophora stylosa, Bruguiera gymnorhiza, and Aegiceras corniculatum were power function models, whereas linear models best fit Kandelia obovata. The carbon density of the artificially planted young mangrove ecosystems in Kaozhouyang Bay was (91.26±17.32) Mg C·hm-2, with a total carbon stock of approximately 35964.65 Mg C. Soil carbon constituted 78.3% to 98.5% of the total carbon stock in these ecosystems. Among the different mangrove communities, vegetation carbon density ranked as follows (from highest to lowest): Aegiceras corniculatum + Kandelia obovata communities, Rhizophora stylosa + Avicennia marina communities, Avicennia marina community, and Bruguiera gymnorhiza community. These results offer valuable insights for assessing carbon storage and guiding ecological restoration efforts in artificially planted mangroves in Guangdong Province and nationwide.
HU Xin , XIONG Lanlan , CHEN Shunyang , ZHANG Huangchen , ZOU Yiyang , ZHANG Jichao , LIU Dongxi , HE Jialu , WU Yuqi , ZHU Zhenjie . Study on biomass models of juvenile mangroves and carbon storage in young mangrove ecosystems*[J]. Journal of Tropical Oceanography, 2025 , 44(4) : 187 -199 . DOI: 10.11978/2024141
表1 红树植物样本基本因子统计量Tab. 1 Basic parameter statistics of mangrove plant samples |
| 树种 | 基径/cm | 株高/m | 树龄/a | 地上生物量/kg | 地下生物量/kg | 地上碳密度系数/% | 地下碳密度系数/% |
|---|---|---|---|---|---|---|---|
| 红海榄 | 1.05~4.14 | 0.46~1.73 | 1~5 | 0.012~1.416 | 0.002~1.069 | 43.79~44.66 | 31.66~40.61 |
| 白骨壤 | 0.70~3.60 | 0.36~1.63 | 1~5 | 0.004~1.531 | 0.001~0.570 | 41.87~44.55 | 35.47~38.79 |
| 桐花树 | 0.60~3.30 | 0.32~0.91 | 1~3 | 0.002~0.326 | 0.001~0.104 | 36.67~40.39 | 35.96~42.20 |
| 木榄 | 0.96~3.76 | 0.32~1.03 | 1~3 | 0.006~0.437 | 0.001~0.131 | 43.06~43.93 | 31.28~37.26 |
| 秋茄 | 0.65~14.32 | 0.36~2.15 | 1~5 | 0.006~3.822 | 0.003~2.215 | 39.28~46.54 | 32.45~40.60 |
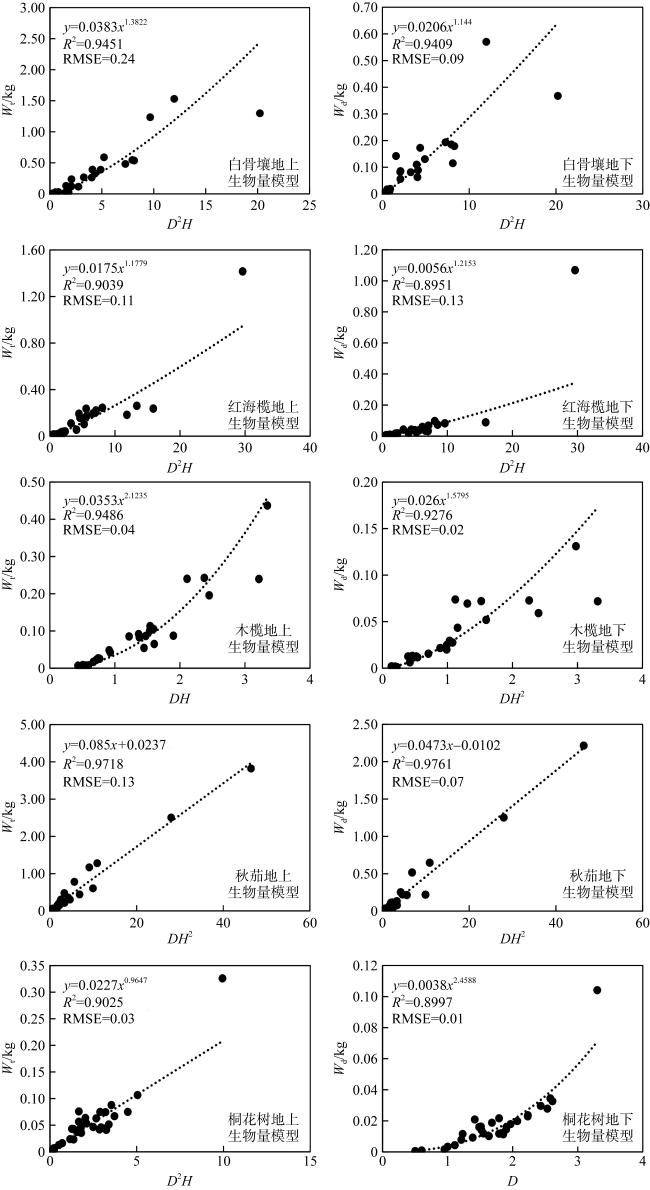
表2 幼龄红树植物生物量模型构建Tab. 2 Biomass model development for juvenile mangrove plants |
| 树种 | 自变量 | 因变量 | R2 | P | 回归模型 | RSS | AIC | RMSE |
|---|---|---|---|---|---|---|---|---|
| 红海榄地上 生物量模型 | D | Wt | 0.88 | <0.01 | Wt = 0.0094×D3.0878 | 0.62 | -114.33 | 0.14 |
| Wt | 0.53 | <0.01 | Wt = 0.2358×D-0.3383 | 0.88 | -103.79 | 0.17 | ||
| H | Wt | 0.61 | <0.01 | Wt = 0.1037×H3.072 | 0.84 | -105.40 | 0.17 | |
| Wt | 0.54 | <0.01 | Wt = 0.6918×H-0.493 | 0.86 | -104.52 | 0.17 | ||
| DH | Wt | 0.88 | <0.01 | Wt = 0.0269×(DH)1.8155 | 0.28 | -135.97 | 0.10 | |
| Wt | 0.77 | <0.01 | Wt = 0.1608×(DH)-0.1843 | 0.44 | -122.53 | 0.12 | ||
| D2H | Wt | 0.90 | <0.01 | Wt = 0.0175×(D2H)1.1779 | 0.23 | -136.10 | 0.11 | |
| Wt | 0.83 | <0.01 | Wt = 0.0388×(D2H)-0.0554 | 0.33 | -131.72 | 0.10 | ||
| DH2 | Wt | 0.82 | <0.01 | Wt = 0.0434×(DH2)1.197 | 0.34 | -130.33 | 0.11 | |
| Wt | 0.89 | <0.01 | Wt = 0.1037×(DH2)-0.0841 | 0.29 | -136.58 | 0.11 | ||
| 红海榄地下 生物量模型 | D | Wd | 0.88 | <0.01 | Wd = 0.0022×D3.4548 | 0.61 | -115.09 | 0.14 |
| Wd | 0.41 | <0.01 | Wd = 0.1651×D-0.2747 | 0.63 | -113.83 | 0.15 | ||
| H | Wd | 0.56 | <0.01 | Wd = 0.0356×H2.4809 | 0.88 | -103.94 | 0.17 | |
| Wd | 0.34 | <0.01 | Wd = 0.3549×H-0.2487 | 0.70 | -110.59 | 0.15 | ||
| DH | Wd | 0.84 | <0.01 | Wd = 0.0097×(DH)1.752 | 0.59 | -113.90 | 0.14 | |
| Wd | 0.62 | <0.01 | Wd = 0.108×(DH)-0.1492 | 0.71 | -124.90 | 0.12 | ||
| D2H | Wd | 0.90 | <0.01 | Wd = 0.0056×(D2H)1.2153 | 0.53 | -146.91 | 0.13 | |
| Wd | 0.76 | <0.01 | Wd = 0.0289×(D2H)-0.0791 | 0.75 | -139.11 | 0.09 | ||
| DH2 | Wd | 0.76 | <0.01 | Wd = 0.0164×(DH2)0.7605 | 0.93 | -100.30 | 0.18 | |
| Wd | 0.78 | <0.01 | Wd = 0.0718×(D2H)-0.0876 | 0.74 | -141.31 | 0.09 | ||
| 白骨壤地上 生物量模型 | D | Wt | 0.92 | <0.01 | Wt = 0.0182×D3.7886 | 2.35 | -74.44 | 0.28 |
| Wt | 0.68 | <0.01 | Wt = 0.4464×D-0.4502 | 1.56 | -86.62 | 0.23 | ||
| H | Wt | 0.74 | <0.01 | Wt = 0.2047×H3.722 | 0.87 | -104.18 | 0.17 | |
| Wt | 0.73 | <0.01 | Wt = 0.9049×H-0.4899 | 1.30 | -92.09 | 0.21 | ||
| DH | Wt | 0.93 | <0.01 | Wt = 0.0595×(DH)2.1007 | 1.12 | -94.54 | 0.19 | |
| Wt | 0.88 | <0.01 | Wt = 0.28×(DH)-0.1703 | 0.59 | -113.80 | 0.14 | ||
| D2H | Wt | 0.95 | <0.01 | Wt = 0.0383×(D2H)1.3822 | 0.71 | -131.98 | 0.14 | |
| Wt | 0.84 | <0.01 | Wt = 0.0844×(D2H)-0.0159 | 0.75 | -106.71 | 0.16 | ||
| DH2 | Wt | 0.89 | <0.01 | Wt = 0.0931×(DH2)1.3826 | 0.63 | -112.00 | 0.14 | |
| Wt | 0.91 | <0.01 | Wt = 0.1705×(DH2)-0.0315 | 0.43 | -123.03 | 0.12 | ||
| 白骨壤地下 生物量模型 | D | Wd | 0.93 | <0.01 | Wd = 0.01×D3.145 | 0.20 | -148.92 | 0.08 |
| Wd | 0.61 | <0.01 | Wd = 0.1179×D-0.0983 | 0.17 | -152.36 | 0.08 | ||
| H | Wd | 0.80 | <0.01 | Wd = 0.1062×H3.4719 | 0.18 | -176.96 | 0.05 | |
| Wd | 0.75 | <0.01 | Wd = 0.2913×H-0.1313 | 0.11 | -165.76 | 0.06 | ||
| DH | Wd | 0.93 | <0.01 | Wd = 0.031×(DH)1.7634 | 0.14 | -156.55 | 0.07 | |
| Wd | 0.79 | <0.01 | Wd = 0.0803×(DH)-0.0281 | 0.10 | -168.47 | 0.06 | ||
| D2H | Wd | 0.94 | <0.01 | Wd = 0.0206×(D2H)1.144 | 0.12 | -163.05 | 0.09 | |
| Wd | 0.74 | <0.01 | Wd =0.0239×(D2H)+0.0099 | 0.12 | -162.04 | 0.06 | ||
| DH2 | Wd | 0.90 | <0.01 | Wd = 0.0473×(DH2)1.1905 | 0.17 | -150.86 | 0.08 | |
| Wd | 0.83 | <0.01 | Wd =0.0526×(D2H)+0.0067 | 0.08 | -175.10 | 0.05 | ||
| 桐花树地上 生物量模型 | D | Wt | 0.90 | <0.01 | Wt = 0.011×D2.3301 | 0.03 | -208.12 | 0.03 |
| Wt | 0.57 | <0.01 | Wt = 0.0704×D-0.0731 | 0.04 | -196.34 | 0.04 | ||
| H | Wt | 0.33 | <0.01 | Wt = 0.1184×H2.1268 | 0.07 | -179.13 | 0.05 | |
| Wt | 0.24 | <0.01 | Wt = 0.1877×H-0.0587 | 0.13 | -161.70 | 0.07 | ||
| 桐花树地上 生物量模型 | DH | Wt | 0.84 | <0.01 | Wt = 0.0378×(DH)1.5403 | 0.02 | -210.19 | 0.03 |
| Wt | 0.67 | <0.01 | Wt = 0.0825×(DH)-0.0402 | 0.03 | -202.75 | 0.03 | ||
| D2H | Wt | 0.90 | <0.01 | Wt = 0.0227×(D2H)0.9647 | 0.02 | -214.66 | 0.03 | |
| Wt | 0.84 | <0.01 | Wt = 0.0275×(D2H)-0.0109 | 0.02 | -203.74 | 0.02 | ||
| DH2 | Wt | 0.71 | <0.01 | Wt = 0.065×(DH2)1.0197 | 0.03 | -199.39 | 0.03 | |
| Wt | 0.68 | <0.01 | Wt = 0.0881×(DH2)-0.0127 | 0.03 | -203.57 | 0.03 | ||
| 桐花树地下 生物量模型 | D | Wd | 0.90 | <0.01 | Wd = 0.0038×D2.4588 | 0.00 | -299.13 | 0.01 |
| Wd | 0.65 | <0.01 | Wd = 0.0241×D-0.0234 | 0.00 | -269.91 | 0.01 | ||
| H | Wd | 0.36 | <0.01 | Wd = 0.0462×H2.2983 | 0.00 | -250.07 | 0.01 | |
| Wd | 0.31 | <0.01 | Wd = 0.0662×H-0.0201 | 0.00 | -249.42 | 0.02 | ||
| DH | Wd | 0.84 | <0.01 | Wd = 0.0143×(DH)1.5553 | 0.00 | -288.83 | 0.01 | |
| Wd | 0.74 | <0.01 | Wd = 0.0272×(DH)-0.0104 | 0.00 | -276.19 | 0.01 | ||
| D2H | Wd | 0.88 | <0.01 | Wd = 0.0084×(D2H)0.9858 | 0.00 | -296.93 | 0.01 | |
| Wd | 0.88 | <0.01 | Wd = 0.009×(D2H)-0.0008 | 0.00 | -299.49 | 0.01 | ||
| DH2 | Wd | 0.70 | <0.01 | Wd = 0.0247×(DH2)1.0295 | 0.00 | -275.08 | 0.01 | |
| Wd | 0.74 | <0.01 | Wd = 0.0293×(D2H)-0.0017 | 0.00 | -276.52 | 0.01 | ||
| 木榄地上 生物量模型 | D | Wt | 0.73 | <0.01 | Wt = 0.0043×D3.6814 | 0.05 | -191.85 | 0.04 |
| Wt | 0.87 | <0.01 | Wt = 0.1475×D-0.1976 | 0.04 | -199.76 | 0.03 | ||
| H | Wt | 0.86 | <0.01 | Wt = 0.2561×H3.4544 | 0.12 | -164.76 | 0.06 | |
| Wt | 0.58 | <0.01 | Wt = 0.3593×H-0.1392 | 0.12 | -163.59 | 0.06 | ||
| DH | Wt | 0.95 | <0.01 | Wt = 0.0353×(DH)2.1235 | 0.04 | -199.35 | 0.04 | |
| Wt | 0.88 | <0.01 | Wt = 0.1151×(DH)-0.0631 | 0.04 | -198.12 | 0.03 | ||
| D2H | Wt | 0.91 | <0.01 | Wt = 0.0158×(D2H)1.413 | 0.06 | -184.64 | 0.04 | |
| Wt | 0.93 | <0.01 | Wt = 0.0333×(D2H)-0.0128 | 0.06 | -200.10 | 0.03 | ||
| DH2 | Wt | 0.95 | <0.01 | Wt = 0.0767×(DH2)1.3609 | 0.06 | -184.47 | 0.04 | |
| Wt | 0.83 | <0.01 | Wt = 0.1054×(DH2)-0.0152 | 0.05 | -188.28 | 0.04 | ||
| 木榄地下 生物量模型 | D | Wd | 0.57 | <0.01 | Wd = 0.0013×D3.8032 | 0.01 | -228.58 | 0.02 |
| Wd | 0.78 | <0.01 | Wd = 0.0452×D-0.0568 | 0.01 | -249.71 | 0.02 | ||
| H | Wd | 0.92 | <0.01 | Wd = 0.1168×H4.2022 | 0.01 | -226.94 | 0.02 | |
| Wd | 0.67 | <0.01 | Wd = 0.1263×H-0.049 | 0.01 | -238.25 | 0.02 | ||
| DH | Wd | 0.88 | <0.01 | Wd = 0.0106×(DH)2.4009 | 0.02 | -216.43 | 0.03 | |
| Wd | 0.84 | <0.01 | Wd = 0.0369×(DH)-0.0177 | 0.00 | -258.62 | 0.01 | ||
| D2H | Wd | 0.80 | <0.01 | Wd = 0.0044×(D2H)1.5536 | 0.02 | -213.67 | 0.03 | |
| Wd | 0.83 | <0.01 | Wd = 0.0102×(D2H)-0.0004 | 0.01 | -255.70 | 0.01 | ||
| DH2 | Wd | 0.93 | <0.01 | Wd = 0.026×(DH2)1.5795 | 0.02 | -251.75 | 0.02 | |
| Wd | 0.80 | <0.01 | Wd =0.0338×(D2H)-0.0025 | 0.01 | -251.02 | 0.01 | ||
| 秋茄地上 生物量模型 | D | Wt | 0.91 | <0.01 | Wt = 0.0243×D2.0144 | 4.83 | -52.82 | 0.40 |
| Wt | 0.82 | <0.01 | Wt = 0.274×D-0.4659 | 3.23 | -64.85 | 0.33 | ||
| H | Wt | 0.87 | <0.01 | Wt = 0.2833×H3.4024 | 6.38 | -44.44 | 0.46 | |
| Wt | 0.66 | <0.01 | Wt = 1.7532×H-1.1884 | 6.51 | -43.82 | 0.47 | ||
| DH | Wt | 0.95 | <0.01 | Wt = 0.0569×(DH)1.3385 | 1.21 | -92.22 | 0.20 | |
| Wt | 0.96 | <0.01 | Wt = 0.1588×(DH)-0.1618 | 0.83 | -103.62 | 0.17 | ||
| D2H | Wt | 0.94 | <0.01 | Wt = 0.04×(D2H)0.8104 | 2.54 | -70.04 | 0.29 | |
| Wt | 0.84 | <0.01 | Wt = 0.011×(D2H)+0.1818 | 3.03 | -64.78 | 0.32 | ||
| DH2 | Wt | 0.95 | <0.01 | Wt = 0.0882×(DH2)0.9796 | 0.58 | -114.47 | 0.14 | |
| Wt | 0.97 | <0.01 | Wt = 0.085×(DH2) + 0.0237 | 0.54 | -116.38 | 0.13 | ||
| 秋茄地下 生物量模型 | D | Wd | 0.95 | <0.01 | Wd = 0.0073×D2.3728 | 4.02 | -58.33 | 0.37 |
| Wd | 0.87 | <0.01 | Wd = 0.1571×D-0.2736 | 0.80 | -106.59 | 0.16 | ||
| H | Wd | 0.85 | <0.01 | Wd = 0.1209×H3.8117 | 2.58 | -71.63 | 0.29 | |
| Wd | 0.60 | <0.01 | Wd = 0.9006×H-0.5968 | 2.38 | -74.00 | 0.28 | ||
| DH | Wd | 0.96 | <0.01 | Wd = 0.0202×(DH)1.5373 | 0.71 | -108.20 | 0.15 | |
| Wd | 0.97 | <0.01 | Wd = 0.0887×(DH)-0.1029 | 0.20 | -146.39 | 0.08 | ||
| D2H | Wd | 0.96 | <0.01 | Wd = 0.0134×(D2H)0.9392 | 1.80 | -80.48 | 0.24 | |
| Wd | 0.89 | <0.01 | Wd = 0.0063×(D2H)-0.0678 | 1.20 | -92.60 | 0.20 | ||
| DH2 | Wd | 0.94 | <0.01 | Wd = 0.0334×(DH2)1.1147 | 0.20 | -146.77 | 0.08 | |
| Wd | 0.98 | <0.01 | Wd = 0.0473×(D2H)-0.0102 | 0.14 | -156.31 | 0.07 |
注: W t和Wd分别表示地上植物生物量和地下植物生物量, 单位为kg; 桐花树生物量为分枝生物量, 单株桐花树生物量应将所有分枝生物量相加计算 |
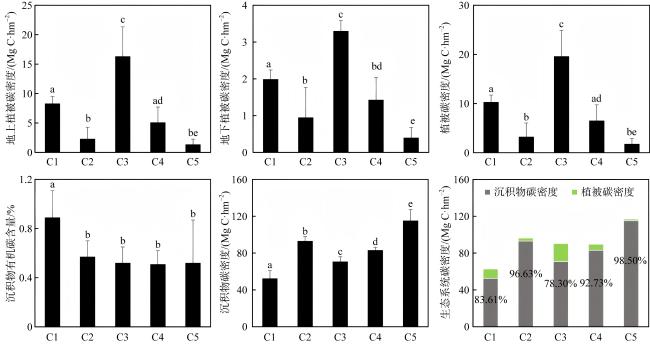
表3 调查站位红树林群落特征Tab. 3 Mangrove community characteristics across sampling stations |
| 站位 | 物种 | 密度/(ind·hm-2) | 基径/cm | 株高/m |
|---|---|---|---|---|
| C1 | 白骨壤 | 11067 | 3.17±0.54 | 1.34±0.26 |
| 红海榄 | 12667 | 2.41±0.58 | 0.70±0.24 | |
| C2 | 木榄 | 10667 | 2.46±1.14 | 1.02±0.37 |
| C3 | 桐花树 | 11200 | 2.43±0.51 | 0.87±0.11 |
| 秋茄 | 3600 | 3.80±2.90 | 0.85±0.45 | |
| C4 | 白骨壤 | 26000 | 2.08±0.80 | 1.09±0.19 |
| C5 | 白骨壤 | 11200 | 2.49±0.38 | 0.77±0.11 |
| 红海榄 | 22000 | 2.09±0.42 | 0.77±0.08 |
| [1] |
董利虎, 李凤日, 2018. 大兴安岭东部主要林分类型乔木层生物量估算模型[J]. 应用生态学报, 29(9): 2825-2834.
|
| [2] |
范航清, 王文卿, 2017. 中国红树林保育的若干重要问题[J]. 厦门大学学报(自然科学版), 56(3): 323-330.
|
| [3] |
高天伦, 管伟, 毛静, 等, 2017. 广东省雷州附城主要红树林群落碳储量及其影响因子[J]. 生态环境学报, 26(6): 985-990.
|
| [4] |
管丽娟, 廖静, 2018. 惠州考洲洋: 用红树林捍卫美丽海湾[J]. 海洋与渔业, (4): 41-42 (in Chinese).
|
| [5] |
何琴飞, 郑威, 黄小荣, 等, 2017. 广西钦州湾红树林碳储量与分配特征[J]. 中南林业科技大学学报, 37(11): 121-126.
|
| [6] |
胡平, 向雪莲, 黄子健, 等, 2024. 外来与乡土红树植物群落生物量动态[J]. 应用与环境生物学报, 30(5): 929-934.
|
| [7] |
胡懿凯, 徐耀文, 薛春泉, 等, 2019. 广东省无瓣海桑和林地土壤碳储量研究[J]. 华南农业大学学报, 40(6): 95-103.
|
| [8] |
黄润霞, 吴卓翎, 彭江炜, 等, 2019. 广东红树植物木榄生物量模型[J]. 西北农林科技大学学报(自然科学版), 47(12): 86-94, 103.
|
| [9] |
金川, 王金旺, 郑坚, 等, 2012. 异速生长法计算秋茄红树林生物量[J]. 生态学报, 32(11): 3414-3422.
|
| [10] |
李海奎, 雷渊才, 曾伟生, 2011. 基于森林清查资料的中国森林植被碳储量[J]. 林业科学, 47(7): 7-12.
|
| [11] |
李林锋, 吴小凤, 刘素青, 2015. 湛江5种红树林树种光合作用特性及光合固碳能力研究[J]. 广西植物, 35(6): 825-832.
|
| [12] |
宁世江, 蒋运生, 邓泽龙, 等, 1996. 广西龙门岛群桐花树天然林生物量的初步研究[J]. 植物生态学报, (1): 57-64.
|
| [13] |
覃国铭, 张靖凡, 周金戈, 等, 2023. 广东省红树林土壤碳储量及固碳潜力研究[J]. 热带地理, 43(1): 23-30.
|
| [14] |
汪珍川, 杜虎, 宋同清, 等, 2015. 广西主要树种(组)异速生长模型及森林生物量特征[J]. 生态学报, 35(13): 4462-4472.
|
| [15] |
王友绍, 2021. 全球气候变化对红树林生态系统的影响、挑战与机遇[J]. 热带海洋学报, 40(3): 1-14.
|
| [16] |
武高洁, 郭志华, 郭菊兰, 等, 2014. 红树林异速生长方程估算生物量研究进展[J]. 湿地科学与管理, 10(3): 61-65.
|
| [17] |
吴雪, 赵鑫, 辜伟芳, 等, 2025. 浙南海岸带人工秋茄红树林与互花米草盐沼土壤碳汇对比研究[J]. 热带海洋学报, 44(1): 1-10.
|
| [18] |
辛琨, 颜葵, 李真, 等, 2014. 海南岛红树林湿地土壤有机碳分布规律及影响因素研究[J]. 土壤学报, 51(5): 1078-1086.
|
| [19] |
鄢春梅, 李文凤, 谢绍茂, 2021. 美丽海湾建设背景下考洲洋海岸带整治与生态修复实践[J]. 广东园林, 43(3): 61-65.
|
| [20] |
姚正阳, 刘建军, 2014. 西安市4种城市绿化灌木单株生物量估算模型[J]. 应用生态学报, 25(1): 111-116.
|
| [21] |
周治刚, 岳文, 李辉权, 等, 2024. 树种类型和潮滩高程对广东湛江高桥红树林碳储量的影响[J]. 热带海洋学报, 43(2): 108-120.
|
| [22] |
朱可峰, 廖宝文, 章家恩, 2011. 广州市南沙红树植物无瓣海桑、木榄人工林生物量的研究[J]. 林业科学研究, 24(4): 531-536.
|
| [23] |
朱耀军, 郭菊兰, 武高洁, 2012. 红树林湿地有机碳研究进展[J]. 生态学杂志, 31(10): 2681-2687.
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}