
Journal of Tropical Oceanography >
Effects of water quality changes on community structures of Symbiodiniaceae and symbiotic bacteria in soft coral Lobophytum sp.
Received date: 2024-07-11
Revised date: 2024-08-02
Online published: 2024-10-14
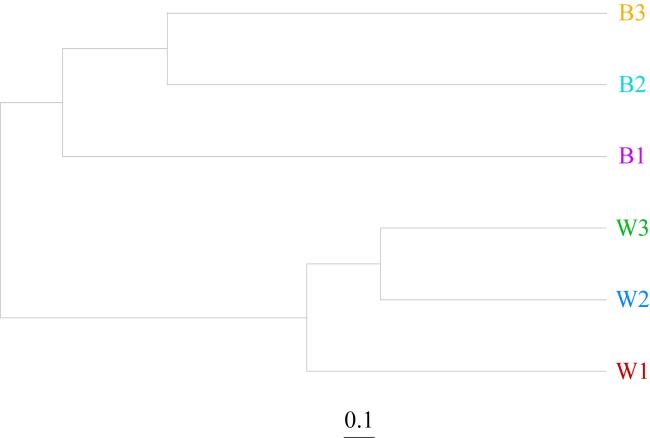
In this study, Lobophytum sp. was cultured under different water qualities for one month, and the community structure of Symbiodiniaceae, ambient water bacteria, and symbiotic bacteria were analyzed using high-throughput sequencing technology. The findings indicated the following. 1) Symbiodiniaceae richness decreased significantly under nutrients conditions ranging from 0 to 80.64 μmol·L-1 nitrate and 0 to 1.05 μmol·L-1 phosphate. Cladocopium sp. dominated the Symbiodiniaceae in all three coral groups, with a relative abundance ranging from 70.25% to 98.13%, exhibiting higher tolerance to low nutrients concentration but greater sensitivity to high nutrients concentration. 2) At the phylum level, water-associated bacteria and coral symbiotic bacteria differed in relative abundance, with all dominant bacterial populations belonging to Proteobacteria, ranging from 45.63% to 86.55% in relative abundance. However, the environmental bacterial diversity at the genus level (Shannon index 4.60~4.97) was higher than that of coral symbiotic bacteria (Shannon index 2.58~3.81), with distinct taxonomic separation between the two communities at the genus level. 3) The coral symbiotic bacterium Cohaesibacter exhibited high tolerance to low nutrient levels, with its relative abundance increasing significantly from < 3% to 40.27% as nutrient levels decreased. Additionally, this genus of soft corals demonstrated strong adaptability to symbiotic bacteria Vibrio, with no significant anomalies observed even at a high Vibrio abundance of 23.71%. These results suggest that the abundance of Symbiodiniaceae and symbiotic bacteria in corals is influenced by ambient water quality, leading to changes in dominant bacteria and alterations in community structure of symbiotic bacteria from corals. Moreover, nutrient fluctuations preferentially shaped the community structure of coral-associated bacteria over environmental bacteria, with more pronounced effects on the former. This study contributes to the growing body of research on soft corals by providing a foundation for understanding how different water quality parameters dynamically affect the structure of symbiotic microorganisms in soft corals. It also offers insights into the effect of water quality fluctuations on soft coral Symbiodiniaceae and bacterial community structure in artificial environments, thereby supporting the development of coral conservation programs.
Key words: Lobophytum; Symbiodiniaceae; symbiotic bacteria; nutrients; community structure
LI Da , WANG Yunzhong , QI Jiguang , YANG Cuihua . Effects of water quality changes on community structures of Symbiodiniaceae and symbiotic bacteria in soft coral Lobophytum sp.[J]. Journal of Tropical Oceanography, 2025 , 44(4) : 136 -144 . DOI: 10.11978/2024138
表1 实验珊瑚缸饲养生物Tab. 1 Marine organisms in experimental coral aquarium |
| 珊瑚缸 | 红皮藻科 | 刺尾鱼科 | 雀鲷科 | 天竺鲷科 | 软珊瑚科 | 海葵科 |
|---|---|---|---|---|---|---|
| 1 | Y | Y | N | N | Y | Y |
| 2 | N | N | Y | Y | N | Y |
| 3 | N | N | N | N | N | Y |
注: Y代表存在, N代表无 |
表2 实验珊瑚缸水质参数Tab. 2 Water quality parameters in experimental coral aquarium |
| 珊瑚缸 | 水温/℃ | 盐度/‰ | pH | 亚硝态氮 /(μmol·L-1) | 氨氮 /(μmol·L-1) | 硝态氮 /(μmol·L-1) | 磷酸盐 /(μmol·L-1) | KH/dKH | 钙离子 /(μmol·L-1) | 镁离子 /(μmol·L-1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 25 | 32 | 8.1~8.4 | 0~0.43 | 0~2.77 | 80.64~161.28 | 1.05~2.11 | 8~9 | 9.48~9.98 | 46.49~52.58 |
| 2 | 25 | 32 | 8.1~8.4 | 0~0.43 | 0~2.77 | 403.20~806.39 | 2.11~5.26 | 8~9 | 9.48~9.98 | 46.49~52.58 |
| 3 | 25 | 32 | 8.1~8.4 | 0~0.43 | 0~2.77 | 32.26~80.64 | 0~1.05 | 8~9 | 9.48~9.98 | 46.49~52.58 |
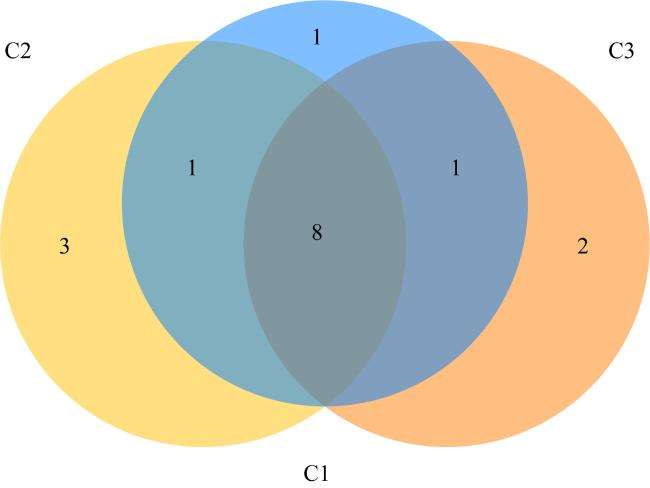
表3 珊瑚共生藻测序数据Tab. 3 Sequencing data of Symbiodiniaceae |
| 样本组名 | 效序列条数 | 平均长度/bp | 最短序 列长度/bp | 最长序 列长度/bp |
|---|---|---|---|---|
| C1 | 106234 | 289.89 | 240 | 369 |
| C2 | 92647 | 305.81 | 201 | 369 |
| C3 | 95787 | 283.77 | 237 | 385 |
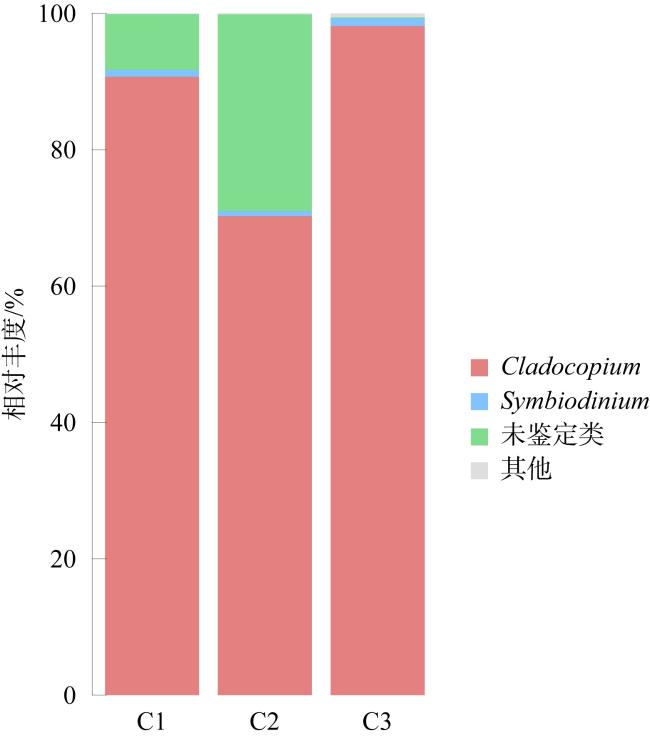
表4 珊瑚共生藻相对丰度情况Tab. 4 Relative abundance of Symbiodiniaceae |
| 样本组名 | Cladocopium 相对丰度/% | Symbiodinium 相对丰度/% | 未鉴定类相 对丰度/% | 其他类 相对丰度/% |
|---|---|---|---|---|
| C1 | 90.69 | 1.01 | 8.18 | 0.12 |
| C2 | 70.25 | 0.75 | 28.85 | 0.15 |
| C3 | 98.13 | 1.14 | 0.13 | 0.60 |
表5 珊瑚共生藻α-多样性指数Tab. 5 α-diversity indices of Symbiodiniaceae |
| 样本组名 | 序列数 | 分类 单元 | Shannon 指数 | Simpson 指数 | Chao 指数 | Ace 指数 | 覆盖率 |
|---|---|---|---|---|---|---|---|
| C1 | 105818 | 20 | 0.42 | 0.82 | 21.00 | 22.82 | 1.00 |
| C2 | 92363 | 22 | 0.74 | 0.56 | 22.00 | 22.00 | 1.00 |
| C3 | 95698 | 14 | 0.15 | 0.95 | 14.00 | 14.00 | 1.00 |
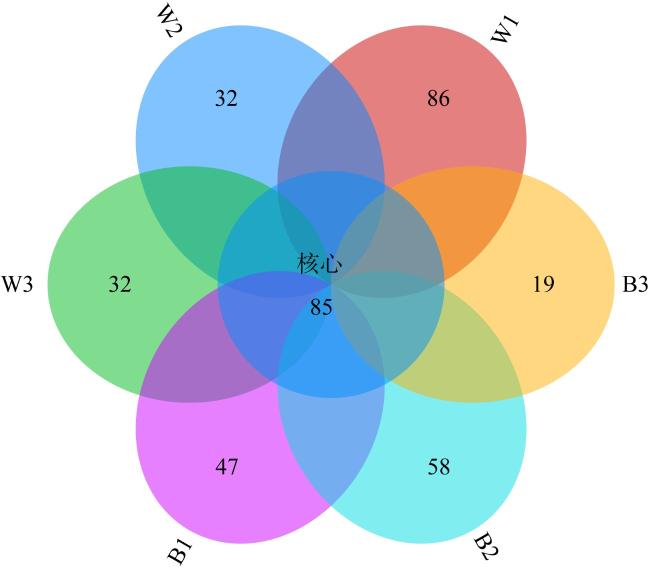
表6 环境水体细菌和珊瑚共生细菌测序数据Tab. 6 Sequencing data statistics for environmental water bacteria and coral symbiotic bacteria |
| 样本组名 | 有效序列条数 | 平均长度/bp | 最短序列长度/bp | 最长序列长度/bp |
|---|---|---|---|---|
| W1 | 70993 | 410.70 | 351 | 468 |
| W2 | 101847 | 410.76 | 350 | 463 |
| W3 | 117782 | 409.87 | 350 | 456 |
| B1 | 53373 | 422.88 | 354 | 462 |
| B2 | 120092 | 416.80 | 350 | 465 |
| B3 | 95859 | 406.93 | 350 | 457 |
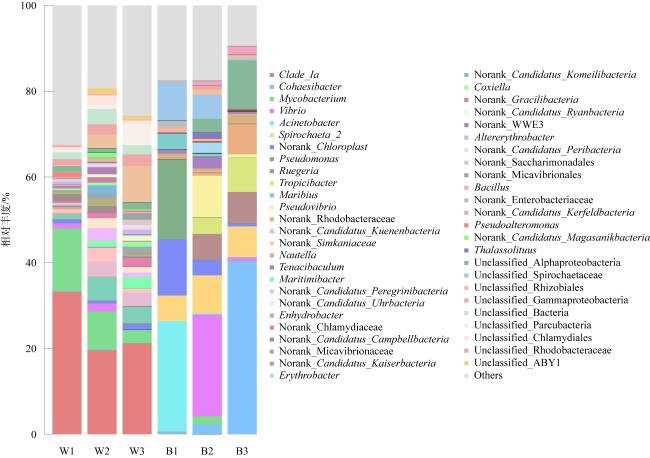
表7 环境水体细菌和珊瑚共生细菌相对丰度(单位: %)Tab. 7 Relative abundance (%) for environmental water bacteria and coral symbiotic bacteria |
| 样本组名 | 变形菌门 | 髌骨菌门 | 放线菌门 | 螺旋体门 | 蓝菌门 | 疣微菌门 | 拟杆菌门 | 蛭弧菌门 | 厚壁菌门 | 浮霉菌门 |
|---|---|---|---|---|---|---|---|---|---|---|
| W1 | 53.46 | 6.21 | 17.33 | 0.01 | 1.52 | 3.62 | 5.68 | 2.17 | 1.15 | 2.26 |
| W2 | 45.63 | 23.51 | 9.98 | 0.02 | 0.83 | 8.37 | 1.74 | 1.53 | 0.38 | 0.80 |
| W3 | 55.98 | 19.64 | 4.21 | 0.05 | 1.55 | 5.47 | 2.22 | 2.96 | 0.14 | 1.16 |
| B1 | 61.03 | 0.10 | 2.03 | 14.93 | 13.07 | 0.38 | 2.43 | 0.01 | 3.61 | 0.77 |
| B2 | 69.75 | 0.07 | 3.24 | 14.56 | 3.52 | 0.64 | 4.73 | 0.26 | 1.90 | 0.31 |
| B3 | 86.55 | 0.51 | 0.42 | 7.60 | 0.64 | 0.49 | 1.21 | 0.88 | 0.57 | 0.44 |
表8 环境水体细菌和珊瑚共生细菌多样性指数Tab. 8 Diversity indices of environmental water bacteria and coral symbiotic bacteria |
| 样本组名 | 序列数 | 分类单元 | Shannon指数 | Simpson指数 | Chao指数 | Ace指数 | 覆盖率 |
|---|---|---|---|---|---|---|---|
| W1 | 53759 | 2746 | 4.60 | 0.13 | 2862.37 | 2924.59 | 0.99 |
| W2 | 70846 | 2658 | 4.97 | 0.05 | 2793.00 | 2887.91 | 0.99 |
| W3 | 112451 | 1809 | 4.89 | 0.06 | 1833.67 | 1822.23 | 1.00 |
| B1 | 47324 | 608 | 3.33 | 0.09 | 647.65 | 656.31 | 1.00 |
| B2 | 116473 | 1007 | 3.81 | 0.06 | 1066.35 | 1052.82 | 1.00 |
| B3 | 88783 | 739 | 2.58 | 0.20 | 803.75 | 829.94 | 1.00 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}