
Journal of Tropical Oceanography >
Study of acute toxicity of Karenia hui on Artemia salina and its hemolytic activity
Copy editor: SUN Cuici
Received date: 2024-10-15
Revised date: 2024-10-23
Online published: 2024-11-05
Supported by
National Key Research and Development Program of China(2022YFC3105201)
Construction of a Phytoplankton Biodiversity Database for Typical Water Sources Based on Environmental DNA Technology
Guangdong Provincial Key R&D Program(2023B1111050011)
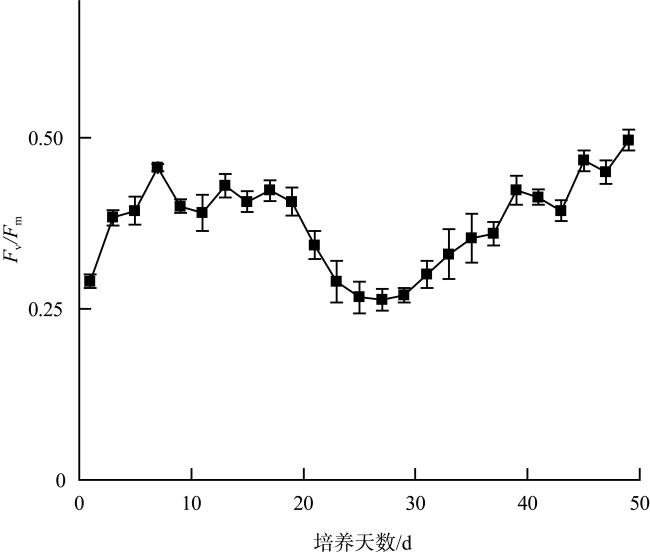
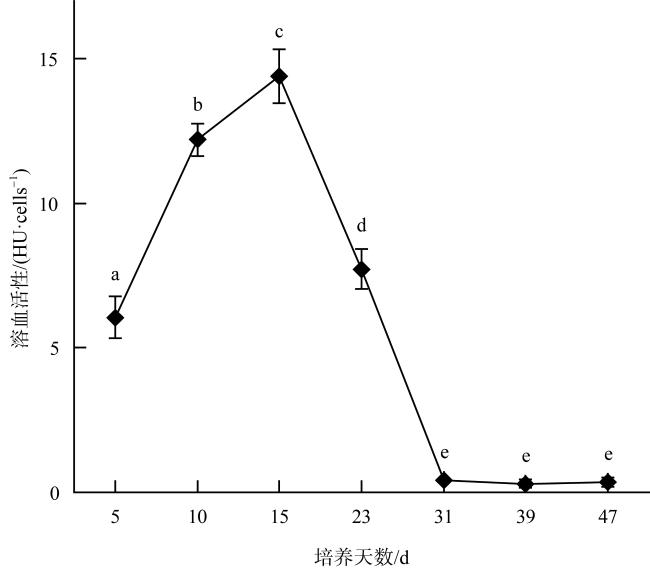
Karenia harmful algal blooms occur frequently worldwide, attracting extensive attention from countries around the globe. Karenia hui, identified as a new species of the genus Karenia in 2024, has been found to be lethal to Oryzias melastigma. Research on this species is currently quite limited. This study investigated its growth characteristics, acute toxicity on Artemia salina, and hemolytic activity at different growth stages. This study aims to reveal the growth curve of K. hui, its toxic effects, and changes in hemolytic activity. The results indicated that K. hui had a unique growth cycle. After the initial exponential growth period and the stable period, it entered the exponential growth period and the stable period again. Karenia hui possessed acute toxicity to Artemia salina, with the death rate of Artemia salina being positively correlated with the density of the algae and increasing with exposure time. In the study of the acute toxicity of K. hui at different growth stages to Artemia salina, the toxicity was the highest on day 23 of the initial stationary phase. The specific death rate of Artemia salina reached its peak at (0.22±0.03)h-1 within 24 hours, and the death rate after 72 hours reached (98.33±4.08)%. The hemolytic activity of K. hui increased during the first exponential growth phase and reached its peak at the end of this phase (14.43×10-6 HU·cells-1). It began to decline during the first stationary phase, and remained low during the second exponential growth phase and stationary phase (0.31×10-6 HU·cells-1). The results of this study can provide a foundation for assessing and mitigating the potential ecological risks posed by K. hui blooms.
Key words: Karenia; Karenia hui; acute toxicity; hemolytic activity
CHEN Shibing , HUANG Bozhu , XIE Xuedong , ZHAO Jinping , LYU Songhui , CEN Jingyi . Study of acute toxicity of Karenia hui on Artemia salina and its hemolytic activity[J]. Journal of Tropical Oceanography, 2025 , 44(1) : 162 -171 . DOI: 10.11978/2024193
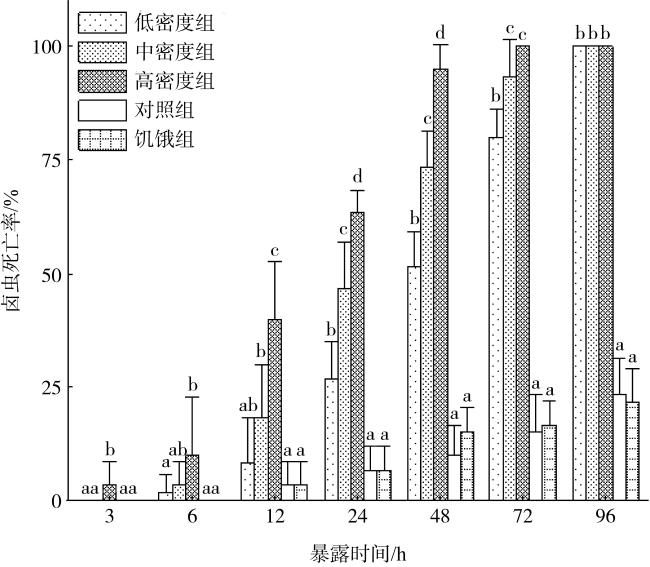
图3 不同密度胡氏凯伦藻暴露下卤虫死亡率不同字母代表同一暴露时间, 不同组之间有显著差异(P<0.05) Fig. 3 The death of Artemia salina under exposure to different concentrations of Karenia hui. Different letters indicate significant differences among different groups at the same exposure time (P<0.05) |
表1 不同密度胡氏凯伦藻暴露下卤虫比死亡速率(h-1)Tab. 1 The specific death rate of Artemia salina under exposure to different concentrations of Karenia hui (h-1) |
| 时间段 | 比死亡速率(h-1) | ||||
|---|---|---|---|---|---|
| 低密度组 | 中密度组 | 高密度组 | 对照组 | 饥饿组 | |
| 0—24h | 0.11±0.03a | 0.19±0.04b | 0.26±0.02c | 0.03±0.02d | 0.03±0.02d |
| 24—48h | 0.10±0.02a | 0.11±0.04a | 0.13±0.03a | 0.01±0.02b | 0.04±0.03b |
| 48—72h | 0.12±0.04a | 0.08±0.04a | 0.02±0.02b | 0.02±0.02b | 0.01±0.01b |
| 72—96h | 0.08±0.03a | 0.03±0.03bc | 0.00±0.00b | 0.04±0.03c | 0.02±0.02bc |
注: 不同字母表示同一暴露时间, 不同组间存在显著差异(P<0.05) |
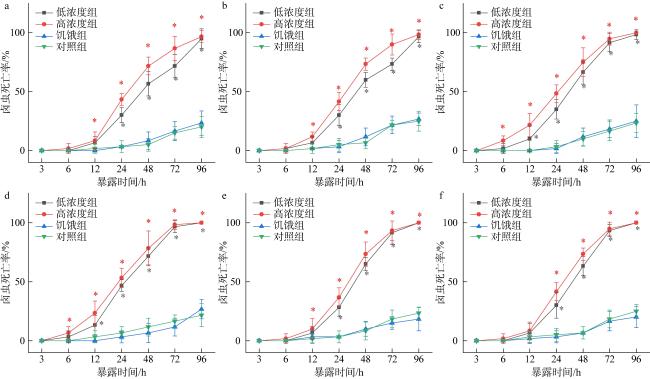
图4 不同生长时期胡氏凯伦藻暴露下卤虫死亡率“*” 表示该组与饥饿组具有显著差异(P<0.05), a—f分别为培养5、10、15、23、31和39d胡氏凯伦藻暴露下卤虫死亡率 Fig. 4 The death rate of Artemia salina exposed to Karenia hui at different growth stages. “*” indicates significant difference between the study group and the control group (P<0.05), a−f represent the death rates of Artemia salina exposed to K. hui on day 5, 10, 15, 23, 31, and 39 of cultivation, respectively |
表2 不同培养天数胡氏凯伦藻暴露下卤虫比死亡速率(h-1)Tab. 2 The specific death rate of Artemia salina exposure to Karenia hui for different culture days (h-1) |
| 时间段 | 密度 组别 | 不同培养天数下的比死亡速率(h-1) | |||||
|---|---|---|---|---|---|---|---|
| 5d | 10d | 15d | 23d | 31d | 39d | ||
| 0—24h | 低 | 0.13±0.03a | 0.13±0.04a | 0.15±0.04a | 0.19±0.02b | 0.12±0.03a | 0.13±0.05a |
| 高 | 0.18±0.02ab | 0.17±0.03ab | 0.20±0.03bc | 0.22±0.03c | 0.15±0.03a | 0.17±0.03ab | |
| 24—48h | 低 | 0.11±0.04a | 0.13±0.06a | 0.13±0.07a | 0.10±0.04a | 0.15±0.03a | 0.14±0.06a |
| 高 | 0.12±0.04a | 0.13±0.03a | 0.11±0.07a | 0.10±0.06a | 0.15±0.03a | 0.13±0.05a | |
| 48—72h | 低 | 0.06±0.06ab | 0.06±0.04a | 0.10±0.02abc | 0.10±0.04abc | 0.11±0.03bc | 0.13±0.05c |
| 高 | 0.06±0.04a | 0.07±0.04a | 0.08±0.07a | 0.08±0.07a | 0.08±0.06a | 0.09±0.03a | |
| 72—96h | 低 | 0.10±0.07a | 0.10±0.03a | 0.03±0.04b | 0.01±0.02b | 0.04±0.04b | 0.03±0.02b |
| 高 | 0.04±0.03a | 0.04±0.04a | 0.02±0.02a | 0.01±0.02c | 0.03±0.03a | 0.02±0.02a | |
注: 不同字母表示同一暴露时间, 不同培养天数组间存在显著差异(P<0.05) |
| [1] |
曹洁茹, 桓清柳, 吴霓, 等, 2015. 光照、温度和氮磷限制对6种典型鱼毒性藻类生长及产毒的影响[J]. 海洋环境科学, 34(3): 321-329.
|
| [2] |
陈宝红, 谢尔艺, 高亚辉, 等, 2015. 米氏凯伦藻对海洋生物致毒作用的研究进展[J]. 福建水产, 37(3): 241-249.
|
| [3] |
崔伟民, 杨维东, 刘洁生, 等, 2009. 米氏凯伦藻溶血毒素的溶血反应特征[J]. 热带亚热带植物学报, 17(3): 237-241.
|
| [4] |
江涛, 吴霓, 钟艳, 等, 2012. 卵圆卡盾藻溶血毒素产毒的影响因素研究[J]. 安全与环境学报, 12(2): 1-5.
|
| [5] |
林雅柔, 岑竞仪, 王建艳, 等, 2020. 中国南海四种凯伦藻种间作用与溶血活性初步研究[J]. 海洋与湖沼, 51(6): 1402-1411.
|
| [6] |
彭喜春, 杨维东, 刘洁生, 等, 2005. 实验室培养球形棕囊藻溶血毒素的提取、分离及其生成特征[J]. 热带亚热带植物学报, 13(1): 25-28.
|
| [7] |
彭颖慧, 刘洁生, 李宏业, 等, 2009. 赤潮藻溶血活性测定标准的建立[J]. 卫生研究, 38(6): 653-656.
|
| [8] |
孙科, 颜天, 周名江, 等, 2010. 米氏凯伦藻对褶皱臂尾轮虫、卤虫和黑褐新糠虾存活的影响[J]. 海洋科学, 34(9): 76-81.
|
| [9] |
王朝晖, 尹伊伟, 齐雨藻, 等, 2001. 珠海桂山岛米氏裸甲藻赤潮对鱼鳃损伤的病理学组织观察[J]. 海洋学报, 23(1): 133-137.
|
| [10] |
严冰, 岑竞仪, 吕颂辉, 2022. 有害甲藻米氏凯伦藻(Karenia mikimotoi)对海洋青鳉鱼(Oryzias melastigm)的急性毒性效应研究[J]. 海洋环境科学, 41(3): 402-407, 415.
|
| [11] |
周成旭, 傅永静, 严小军, 2007. 4种典型有害赤潮原因种的溶血特性研究[J]. 生态毒理学报, 2(1): 78-82.
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}