
Journal of Tropical Oceanography >
Morphological characteristics of epilithic algal matrix and their spatial and temporal patterns in a typical fringing reef of Sanya
Received date: 2024-11-21
Revised date: 2024-12-08
Online published: 2025-01-13
Supported by
National Science & Technology Fundamental Resources Investigation Program of China(2022FY100602)
National Natural Science Foundation of China(42176118)
Science and Technology Planning Project of Guangdong Province, China(2023B1212060047)
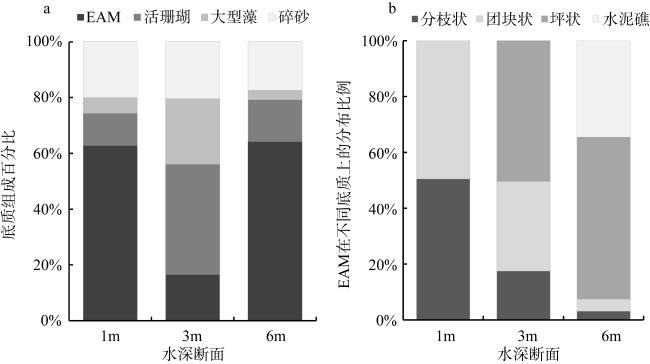
The epilithic algal matrix (EAM) is widely distributed in coral reef ecosystems and plays a crucial role in key processes such as primary production, nutrient recycling, sediment deposition, and coral reef phase shifts. To investigate the morphological characteristics and distribution patterns of EAM, a survey was conducted in a typical Sanya fringing reef in January, April, July, and October 2022, covering three transects (1 m, 3 m, 6 m). Substrate types and their coverage were recorded via video, and three substrate types (branched, massive, planiform) with epiphytic algae were sampled by scuba diving. The results revealed significant spatial but no seasonal differences in EAM coverage in the study area. Coverage was higher at 1 m and 6 m depths (66.96%) but lower at 3 m (16.55%), showing a negative correlation with live coral coverage. EAM primarily colonized highly porous hard substrates, with the highest coverage (98%) observed on dead planiform coral reefs. However, dead massive coral reefs exhibited the highest algal height [(11.16 ± 0.68) mm], highest biomass [(118.51±33.64) g∙m-2], and greatest organic matter content [(102.49±32.94) g C∙m-2], likely due to their higher surface porosity. EAM morphological characteristics and biomass varied with depth and season, peaking at 3 m and reaching their lowest values at 1 m. Temporally, EAM height, density, and biomass were highest in summer and lowest in winter. The study indicates that the benthic community in Sanya’s Luhuitou area is dominated by long sediment-laden algal turfs (LSATs) with high sediment content and heights exceeding 5 mm. This community type is unfavorable for coral larvae sediment and is avoided by herbivorous fish. EAM morphological traits and organic matter content exhibit distinct variations across substrates, depths, and seasons. Additionally, the rich organic matter in EAM may serve as a potential food source for benthic invertebrates, playing a vital role in the coral reef ecosystem’s food web.
HU Simin , ZHOU Tiancheng , ZHANG Chen , JIA Nan , LIU Sheng , LI Tao , HUANG Hui . Morphological characteristics of epilithic algal matrix and their spatial and temporal patterns in a typical fringing reef of Sanya[J]. Journal of Tropical Oceanography, 2025 , 44(4) : 95 -105 . DOI: 10.11978/2024212
表 1 研究区域环境因子Tab. 1 Environmental parameters of the study area |
| 季节 | 水深/m | 温度/℃ | 盐度/‰ | 悬浮物/(mg·L-1) | 溶解无机氮/(μmol·L-1) | 溶解无机磷/(μmol·L-1) | 硅酸盐/(μmol·L-1) |
|---|---|---|---|---|---|---|---|
| 冬 | 1 | 24.25±0.02 | 33.00±0.03 | 4.43±1.61 | 0.12±0.17 | 0.18±0.01 | 8.13±0.88 |
| 3 | 24.21±0.01 | 33.02±0.00 | 1.30±0.54 | 0.11±0.09 | 0.18±0.01 | 5.41±0.31 | |
| 6 | 24.19±0.00 | 33.02±0.00 | 3.40±1.13 | 0.66±0.14 | 0.31±0.21 | 5.55±0.80 | |
| 春 | 1 | 27.23±0.01 | 33.66±0.00 | 4.44±2.29 | 1.12±0.23 | 0.03±0.01 | 6.26±0.42 |
| 3 | 27.08±0.02 | 33.65±0.00 | 3.74±0.67 | 1.52±0.07 | 0.02±0.00 | 5.77±0.09 | |
| 6 | 27.07±0.00 | 33.66±0.00 | 2.75±1.81 | 2.16±0.17 | 0.02±0.00 | 6.47±0.14 | |
| 夏 | 1 | 27.84±0.04 | 33.55±0.01 | 12.93±0.39 | 0.79±0.05 | 0.01±0.00 | 7.97±0.21 |
| 3 | 27.31±0.14 | 33.57±0.02 | 12.80±0.78 | 0.60±0.08 | 0.01±0.00 | 5.92±0.08 | |
| 6 | 26.14±0.15 | 33.56±0.59 | 14.62±0.30 | 0.88±0.29 | 0.01±0.00 | 8.50±0.18 | |
| 秋 | 1 | 27.10±0.02 | 32.06±0.01 | 4.67±1.73 | 1.16±0.28 | 0.05±0.01 | 5.38±1.46 |
| 3 | 27.08±0.01 | 32.05±0.00 | 2.22±0.41 | 1.72±0.12 | 0.03±0.03 | 7.46±0.26 | |
| 6 | 27.10±0.00 | 32.17±0.00 | 2.60±0.82 | 1.75±0.04 | 0.02±0.01 | 8.45±0.45 |
表 2 不同类型底质的表观特征Tab. 2 Classification and apparent characteristics of different substrate types |
| 底质生境 | 简称 | 描述 | n | 孔隙度/% | 表面积/m2 | 藻席覆盖度/% |
|---|---|---|---|---|---|---|
| 分枝状珊瑚礁石 | F | 死亡的分枝状珊瑚骨骼形成的坚硬、多孔基底, 主要位于1m、3m水深 | 60 | 7.71±2.38 | 0.02±0.01 | 32.64±12.13 |
| 团块状珊瑚礁石 | T | 死亡的团块状珊瑚骨骼形成的坚硬、多孔基底, 在各水深都有分布 | 81 | 3.05±0.05 | 0.28±0.20 | 65.13±25.39 |
| 坪状珊瑚礁石 | P | 死亡的坪状珊瑚骨骼形成的坚硬、多孔基底, 在各水深都有分布 | 64 | 9.93±4.97 | 9.90±14.07 | 98.07±3.25 |
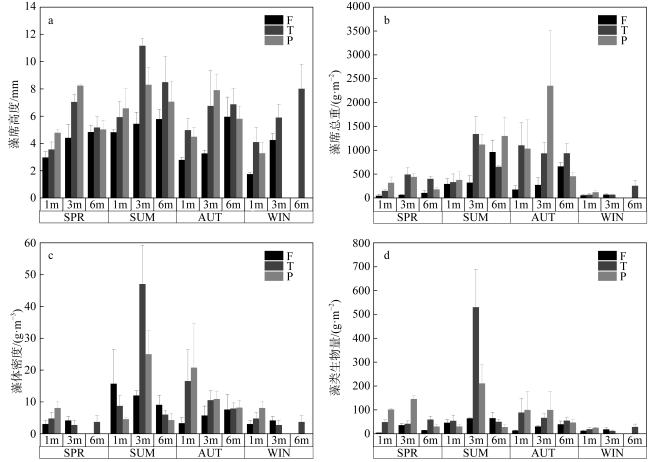
图2 不同季节和水深礁石表生藻席的表观特征差异a. 藻席高度; b. 藻席总重; c. 藻体密度; d. 藻类生物量; F: 分枝状珊瑚断枝; T: 团块状珊瑚礁石; P: 坪状珊瑚礁石; WIN: 冬季; SPR: 春季; SUM: 夏季; AUT: 秋季 Fig. 2 Apparent characteristics of EAM on reef substrates across different seasons and water depths. (a) Matrix height; (b) total matrix weight; (c) algal density; d. algal biomass). F: Branched dead coral; T: massive dead coral; P: planiform dead coral; WIN: winter; SPR: spring; SUM: summer; AUT: autumn |
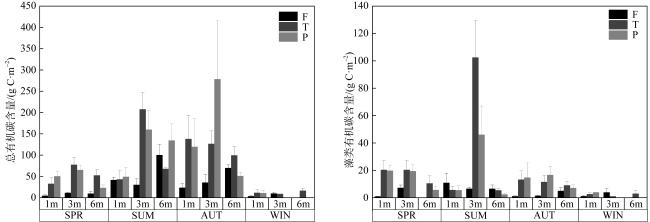
图3 不同季节和水深礁石表生藻席的有机碳储量(a. 总有机碳含量; b. 藻类有机碳含量)F: 分枝状珊瑚断枝; T: 团块状珊瑚礁石; P: 坪状珊瑚礁石; WIN: 冬季; SPR:春季; SUM:夏季; AUT:秋季) Fig. 3 Organic carbon storage in EAM of reef rocks across different seasons and water depths (a. total organic carbon content; b. algal organic carbon content). F: Branched dead coral; T: massive dead coral; P: planiform dead coral; WIN: winter; SPR: spring; SUM: summer; AUT: autumn |
表 3 环境因子与礁石表生藻席特征的相关性系数Tab. 3 Correlation coefficients between environmental factors and feature parameters of EAM |
| 环境因子 | 藻席高度 | 藻席总重 | 藻体密度 | 总有机碳 | 藻类生物量 | 沉积物负荷 | 藻类有机碳 | 藻类有机碳比例% |
|---|---|---|---|---|---|---|---|---|
| 水深 | 0.440** | 0.178* | −0.141 | 0.099 | −0.023 | 0.192* | −0.055 | −0.322** |
| 孔隙度 | −0.144 | 0.131 | 0.029 | 0.056 | −0.048 | 0.146 | −0.107 | 0.001 |
| 温度 | 0.171 | 0.323** | 0.372** | 0.302** | 0.295** | 0.294** | 0.278** | −0.169 |
| 盐度 | 0.139 | −0.246** | 0.077 | −0.229* | 0.112 | −0.279** | 0.166 | 0.320** |
| 悬浮物 | 0.286** | 0.207* | 0.272** | 0.179 | 0.306** | 0.169 | 0.235* | −0.075 |
| 溶解无机氮 | 0.157 | 0.233* | 0.025 | 0.158 | −0.006 | 0.248** | −0.052 | −0.281** |
| 溶解无机磷 | −0.116 | −0.309** | −0.274** | −0.319** | −0.219* | −0.292** | −0.210* | 0.154 |
| 硅酸盐 | 0.016 | 0.144 | −0.107 | 0.080 | −0.104 | 0.169 | −0.186 | −0.137 |
注: n=123; 粗体表示差异显著,其中*表示P<0.05, **表示P<0.01 |
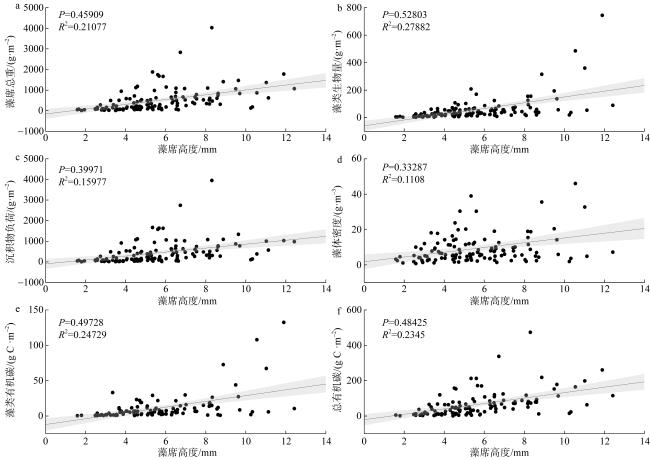
图4 藻席高度与藻席其他特征的相关性分析a—f分别为藻席高度与藻席总重、藻类生物量、沉积物负荷、藻体密度、藻类有机碳、总有机碳的散点图; 趋势线两侧条带表示95%置信区间 Fig. 4 Correlation analysis between EAM height and other characteristics of EAM. Panels a-f are scatter plots of algal matrix height versus total matrix weight, algal biomass, sediment load, algal density, algal organic carbon, and total organic carbon, respectively; the bands on either side of the trend line represent the 95% confidence interval |
| [1] |
梁宇娴, 俞晓磊, 郭亚娟, 等, 2020. 3种传统方法对不同珊瑚表面积测量的适用性及其校准方法—以3D扫描技术为基准[J]. 热带海洋学报, 39(1): 85-93.
|
| [2] |
廖芝衡, 2021. 南海珊瑚群落和底栖海藻的空间分布特征及其生态影响[D]. 南宁: 广西大学.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}