
Journal of Tropical Oceanography >
Community structure of rhizosphere eukaryotic microorganisms and its influencing factors of three seagrass species along the coast of Shandong Province
Received date: 2025-01-02
Revised date: 2025-01-24
Online published: 2025-02-26
Supported by
National Natural Science Foundation of China(41976147)
Natural Science Foundation of Shandong Province(ZR2024MD008)
Seagrass beds, mangroves, and coral reefs are known as the three typical offshore marine ecosystems on Earth. Seagrass beds, which possess extremely high ecological service functions, are hailed as the “lungs of the ocean”. Seagrasses have a relatively high level of primary productivity and play a core role in maintaining the balance and stability of the marine ecosystem. Rhizosphere eukaryotes are crucial for seagrass growth and development, playing a significant role in sustaining seagrass health and facilitating the cycling of nutrients such as carbon and nitrogen within the ecosystem. In this study, high-throughput sequencing technology was employed to investigate the community structure and diversity of eukaryotes in the rhizosphere of Ruppia sinensis, Zostera marina, and Zostera japonica along the coastal areas of Shandong Province. The results showed that seagrass-associated eukaryotes were mainly composed of algae, metazoans, and protozoans, with Chlorophyta and Annelida being the dominant groups. The community structure of eukaryotes in the seagrass rhizosphere was significantly correlated with the total carbon content, nitrite content, ammonium content, and sediment particle size. Co-occurrence network analysis further revealed that the interactions among eukaryotes in Zostera japonica were more complex. This study elucidates the community structure and influencing factors of eukaryotes in the seagrass rhizosphere, laying a theoretical foundation for further exploration of nutrient cycling and energy flow in seagrass ecosystems.
YU Zhen , GUO Xiangrui , LIU Xuerui , SUN Hao , ZHANG Yanying . Community structure of rhizosphere eukaryotic microorganisms and its influencing factors of three seagrass species along the coast of Shandong Province[J]. Journal of Tropical Oceanography, 2025 , 44(5) : 86 -96 . DOI: 10.11978/2025003
表1 海草根际真核生物α多样性指数分析Tab. 1 Analysis of α diversity indices of eukaryotic microorganisms in the rhizosphere of seagrasses |
| 多样性指数 | 中国川蔓草 | 鳗草 | 日本鳗草 | P |
|---|---|---|---|---|
| Ace指数 | 243.50±93.61 | 321.50±196.5 | 321.80±138.10 | 0.7351 |
| Chao指数 | 235.10±92.07 | 319.90±191.90 | 316.90±138.30 | 0.7351 |
| 香农指数 | 2.92±0.8002 | 3.07±1.18 | 2.15±1.31 | 0.5950 |
| 辛普森指数 | 0.13±0.11 | 0.21±0.19 | 0.41±0.31 | 0.1988 |
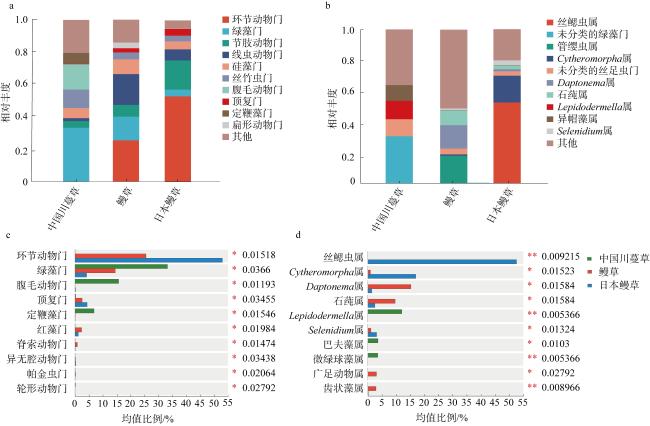
图4 中国川蔓草、鳗草和日本鳗草根际沉积物真核生物群落结构及主要类群的相对丰度组间差异分析a. 门水平的群落组成; b. 属水平的群落组成; c. 门水平的组间差异分析; d. 属水平的组间差异分析 Fig. 4 Analysis of eukaryotic community composition and relative abundance of major groups in rhizosphere sediments of Ruppia sinensis, Zostera marina and Zostera japonica. (a) Community composition at phylum level; (b) community composition at genus level; (c) intergroup difference analysis at phylum level; (d) intergroup difference analysis at genus level |
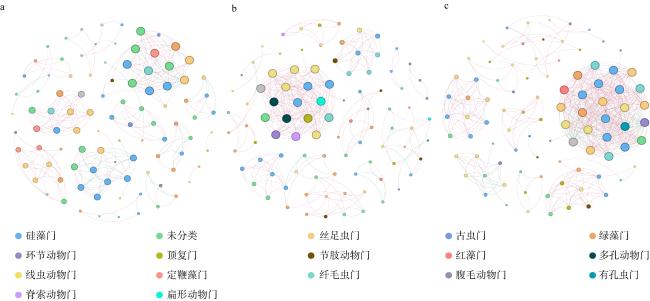
图5 3种海草根际真核生物群落在ASV水平上的共现性网络图a. 中国川蔓草; b. 鳗草; c. 日本鳗草。 红色线表示正相关关系, 绿色线表示负相关关系 Fig. 5 Co-occurrence network diagrams of eukaryotic communities of different seagrass at ASV level. (a) Ruppia sinensis; (b) Zostera marina; (c) Zostera japonica. Red lines indicate positive correlations, green lines indicate negative correlations |
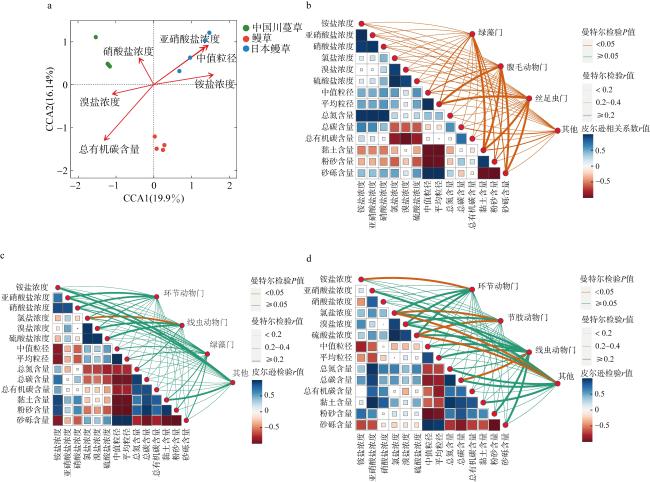
图6 海草根际真核生物群落与环境因子之间的关系a. 海草根际真核生物与环境因子的CCA分析; b、c、d分别为中国川蔓草、鳗草和日本鳗草主要真核生物类群与环境因子的相关性分析 Fig. 6 Relationships between seagrass rhizosphere eukaryotic microbial communities and environmental factors. (a) CCA analysis of seagrass rhizosphere eukaryotic microorganisms and environmental factors; (b), (c), and (d) are the correlation coefficients between the environmental factors and major eukaryotic species in the rhizosphere of Ruppia sinensis, Zostera marina and Zostera japonica |
| [1] |
凌娟, 梁童茵, 岳维忠, 等, 2023. 热带海草泰来草沉积物真菌的群落结构、功能与分子生态网络研究[J]. 热带海洋学报, 42(5): 64-75.
|
| [2] |
刘鹏远, 2022. 黄渤海日本鳗草沉积物三域微生物的分布特征、生态功能及驱动因素[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所).
|
| [3] |
刘鹏远, 张海坤, 陈琳, 等, 2019. 黄渤海海草分布区日本鳗草根际微生物群落结构特征及其功能分析[J]. 微生物学报, 59(8): 1484-1499.
|
| [4] |
宋增磊, 2023. 微生物在黄河三角洲互花米草与日本鳗草生态竞争中的作用及机制分析[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所).
|
| [5] |
孙延瑜, 2021. 威海泻湖海草床共附生微生物群落结构特征及硫入侵过程[D]. 烟台: 中国科学院大学(中国科学院烟台海岸带研究所).
|
| [6] |
孙延瑜, 宋增磊, 刘鹏远, 等, 2021. 威海天鹅湖大叶藻(Zostera marina)与日本鳗草(Zostera japonica)根际微生物群落结构及其驱动机制[J]. 微生物学报, 61(9): 2675-2692.
|
| [7] |
王天雨, 2023. 山东东部沿海两种海草根际微生物群落结构及潜在功能[D]. 烟台: 烟台大学.
|
| [8] |
徐少春, 2021. 黄渤海鳗草(Zostera marina)种群特征及生态修复研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所).
|
| [9] |
薛曌, 王兰, 孟华旦尚, 等, 2023. 色林错表层水体真核微生物多样性和群落分布格局[J]. 冰川冻土, 45(5): 1652-1666.
|
| [10] |
于培, 2017. 鳗草对沉积物微生物群落的影响及其氮代谢的初步研究[D]. 济南: 山东大学.
|
| [11] |
张晓珂, 梁文举, 李琪, 2018. 我国土壤线虫生态学研究进展和展望[J]. 生物多样性, 26(10): 1060-1073.
|
| [12] |
周毅, 江志坚, 邱广龙, 等, 2023. 中国海草资源分布现状、退化原因与保护对策[J]. 海洋与湖沼, 54(5): 1248-1257.
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}