
Journal of Tropical Oceanography >
Ecological and environmental effects of boiled and inactivated remains of crown-of-thorns starfish
Received date: 2024-12-16
Revised date: 2025-02-20
Online published: 2025-03-04
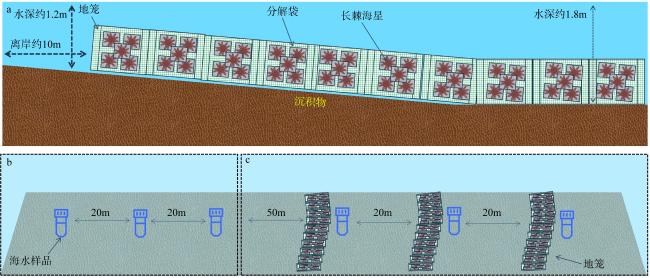
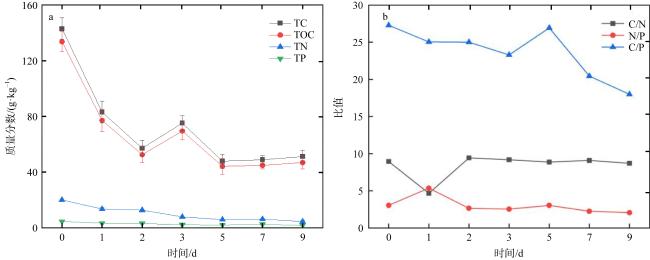
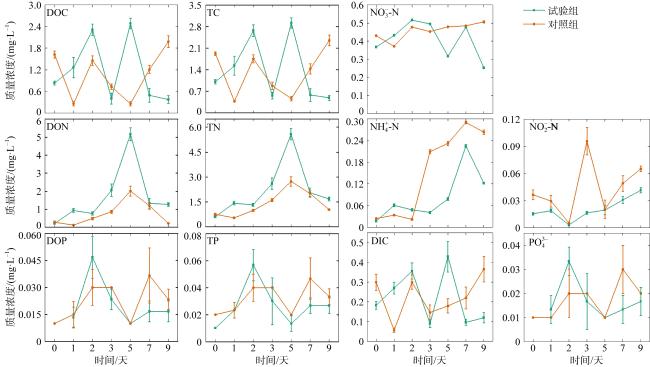
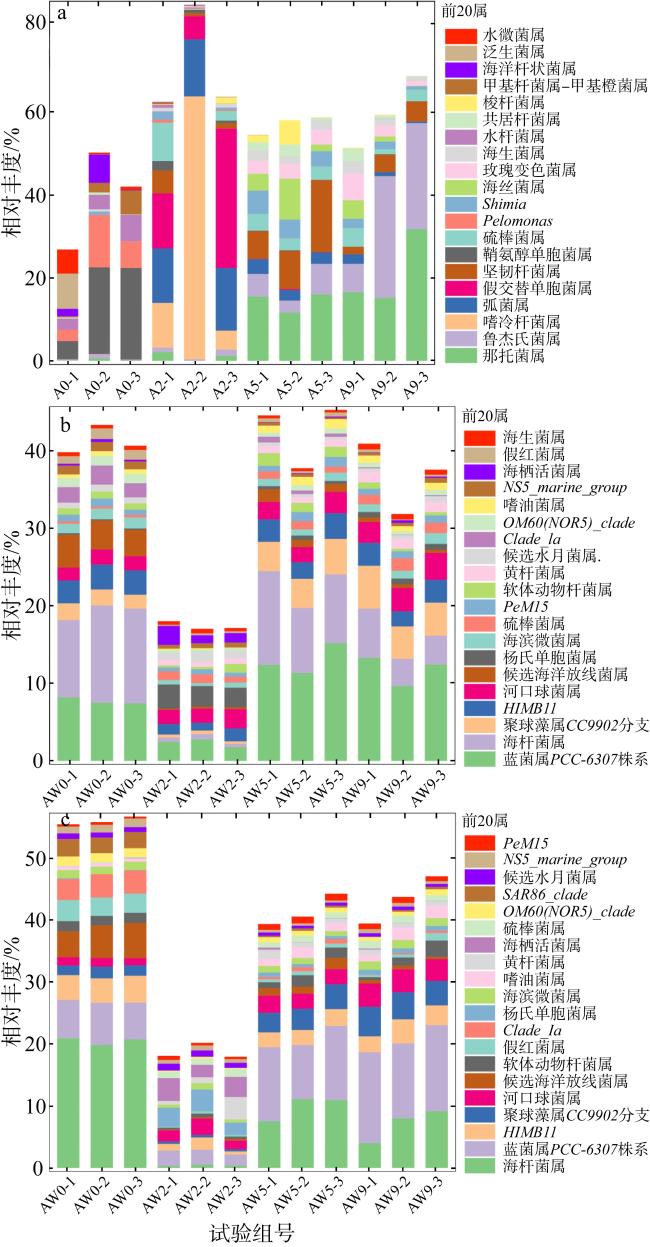
The outbreak of crown-of-thorns starfish (Acanthaster planci, CoTS) poses a significant threat to the health of coral reef ecosystems. Artificial removal is considered one of the most practical and effective methods for addressing local outbreaks of CoTS. However, the marine environmental impact of discarding inactivated CoTS after capture is currently unclear. In this study, an in situ experiment was conducted to assess the ecological and environmental effects of boiled and inactivated CoTS remains. The results showed that boiled CoTS tissues decomposed within 2 days, with the skeletal remains breaking down into granular fragments. By the 9th day after returning the CoTS to the sea, 63.20% of carbon, 62.18% of nitrogen, and 44.17% of phosphorus were released into the water, resulting in an increase of (0.08 ± 0.06) mg·L-1 in carbon, (0.08 ± 0.08) mg·L-1 in nitrogen, and a decrease of 0.01 mg·L-1 in phosphorus concentrations. In addition, the dominant bacteria on the surface of the inactivated CoTS primarily belonged to Proteobacteria, Cyanobacteria, Actinobacteria, and Bacteroidetes. Dominant genera included Sphingomonas (Bacteroidetes), and Ruegeria, Pelomonas, Nautella, and Tenacibaculum (Proteobacteria), which are associated with CoTS decomposition. The boiled and inactivated CoTS remains decomposed rapidly and released nutrients directly. A small amount of inactivated CoTS does not cause significant adverse environmental effects, suggesting this method is a relatively economical and eco-friendly approach for managing CoTS outbreaks.
Key words: crown-of-thorns starfish; outbreak; coral; marine environment; ecological effect
LIU Jiangen , LUO Hongtian . Ecological and environmental effects of boiled and inactivated remains of crown-of-thorns starfish[J]. Journal of Tropical Oceanography, 2025 , 44(5) : 189 -200 . DOI: 10.11978/2024234
图2 长棘海星(长棘海星试验组)分解过程中的形态变化a. 第0天; b. 第1天; c. 第2天; d. 第3天; e. 第5天; f. 第7天; g. 第9天 Fig. 2 Morphological changes during the decomposition process of CoTS |
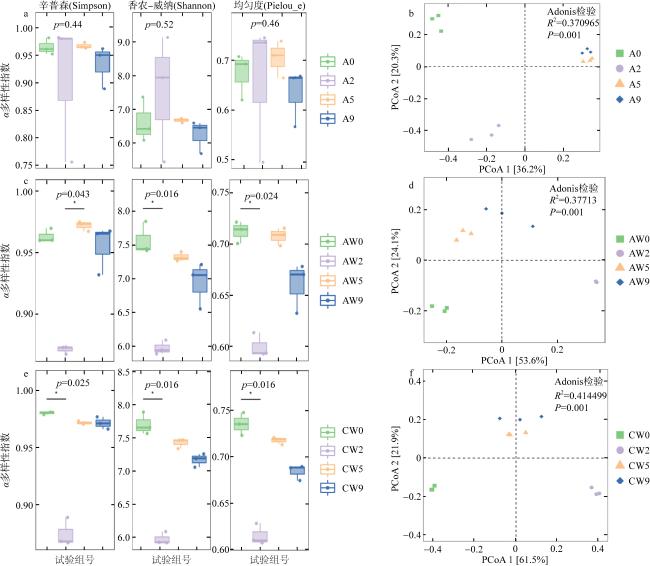
图5 长棘海星分解过程中的细菌多样性a. 海星组; b. 试验组; c. 对照组 Fig. 5 Bacterial diversity during CoTS decomposition process |
| [1] |
康霖, 2016. 西沙群岛海洋渔业资源调查研究[J]. 海洋与渔业 (2): 64-66.
|
| [2] |
李元超, 吴钟解, 梁计林, 等, 2019. 近15年西沙群岛长棘海星爆发周期及爆发原因分析[J]. 科学通报, 64(33): 3478-3484.
|
| [3] |
马清霞, 李宁, 李学刚, 等, 2012. 模拟研究沙海蜇消亡过程中海水pH变化及对海水酸化的影响[J]. 海洋科学, 36(12): 12-18.
|
| [4] |
曲长凤, 宋金明, 李宁, 等, 2016. 海水中沙海蜇消亡对水体碳、氮、磷的释放与补充[J]. 应用生态学报, 27(1): 299-306.
|
| [5] |
宋金明, 李鹏程, 1996. 南沙群岛海域沉积物-海水界面间营养物质的扩散通量[J]. 海洋科学, 20(5): 43-50.
|
| [6] |
夏荣林, 宁志铭, 余克服, 等, 2022. 长棘海星爆发对珊瑚礁区沉积物营养盐动力学的影响研究[J]. 海洋学报, 44(8): 23-30.
|
| [7] |
姚秋翠, 余克服, 廖芝衡, 等, 2022. 棘冠海星爆发及其对珊瑚礁的生态影响研究进展[J]. 生态学报, 42(18): 7517-7528.
|
| [8] |
朱士兵, 胡丹妮, 张会领, 等, 2019. 海口湾中间岸段海滩剖面短期时空变化及沉积动态分析[J]. 热带海洋学报, 38(5): 77-85.
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}