
Journal of Tropical Oceanography >
Virtual screening of potential anti-osteoarthritic peptides from sea cucumber and their mechanisms based on network pharmacology, molecular docking, and quantum chemistry calculation*
Received date: 2025-01-15
Revised date: 2025-03-26
Online published: 2025-04-01
Supported by
Marine Economic Development Project of Guangdong(GDNRC[2024]49)
Osteoarthritis (OA) is a chronic degenerative joint disease characterized by the progressive loss of articular cartilage and the destruction of joint structures. This study investigates the potential role of sea cucumber peptides in combating OA. Potential bioactive peptides from sea cucumber protein were identified through computer simulations of gastrointestinal digestion and online database predictions. Using network pharmacology, molecular docking, and quantum chemical calculations, a sea cucumber peptide candidate (phenylalanine-aspartic acid-proline-valine-isoleucine-glutamic acid-glutamic acid-tyrosine-histidine-asparagine-glycine-phenylalanine, FDPVIEEYHNGF) with strong anti-OA activity was virtually screened. Further analysis suggests that this peptide may alleviate OA by modulating the IL-17 and TNF signaling pathways, inhibiting inflammation, collagen degradation, and the activity of matrix metalloproteinases (MMPs). The sea cucumber peptide may form hydrogen bonds with MMP9 and IL-17RA, potential core targets for OA, thereby influencing the related signaling pathways, reducing inflammation, intervening in extracellular matrix remodeling, and mitigating collagen degradation. This study provides new insights into the application of sea cucumber peptides as functional food ingredients in the development of therapeutic foods for OA.
DUAN Ailing , LI Si , ZHAO Xiangtan , CHEN Hua , WAN Peng , CHEN Deke , CAI Bingna , PAN Jianyu . Virtual screening of potential anti-osteoarthritic peptides from sea cucumber and their mechanisms based on network pharmacology, molecular docking, and quantum chemistry calculation*[J]. Journal of Tropical Oceanography, 2025 , 44(6) : 176 -187 . DOI: 10.11978/2025011
表1 选取的海参蛋白信息Tab. 1 Information of selected sea cucumber proteins |
| UniProt检索号 | 蛋白检索名 | 蛋白名称 | 氨基酸总数 | 疏水性氨基酸比例 | 支链氨基酸比例 |
|---|---|---|---|---|---|
| Q868M7 | CEL3_CUCEC | Galactose/N-acetylgalactosamine-binding lectin CEL-III, Gal/GalNAc-binding lectin CEL-III, CEL-III, Hemolytic lectin CEL-III, Lectin CEL-III | 442 | 55.66% | 17.65% |
| A0A286QZ36 | RAC1_STIJA | Ras-related protein Rac1, AjRac1 | 192 | 55.73% | 23.44% |
| P80018 | GLBC_MOLAR | Globin C, coelomic | 159 | 55.35% | 22.01% |
| P48557 | H2B_HOLTU | Histone H2B | 123 | 51.22% | 15.45% |
| P15161 | GLB1_PARCH | Globin-1, Globin I | 158 | 55.70% | 22.15% |
| P80017 | GLBD_MOLAR | Globin D, coelomic | 159 | 61.64% | 12.58% |
| P21251 | CALM_STIJA | Calmodulin, CaM | 149 | 52.35% | 16.11% |
| Q9XY07 | KARG_STIJA | Arginine kinase, AK, EC:2.7.3.3 | 370 | 50.81% | 19.73% |
| P62776 | H4_HOLTU | Histone H4 | 103 | 85.44% | 70.87% |
| P28766 | SODM_APOCL | Superoxide dismutase [Mn], mitochondrial, EC:1.15.1.1 | 144 | 69.44% | 20.14% |
| P14309 | PHI0_HOLTU | Sperm-specific protein Phi-0 | 78 | 46.15% | 6.41% |
| Q7M3Y0 | SJL1_STIJA | Lectin 1, Stichopus japonicas lectin 1, SjL-1 | 143 | 60.84% | 11.19% |
| P19477 | FIBA_APOPA | Fibrinogen-like protein A, FREP-A | 282 | 48.94% | 16.67% |
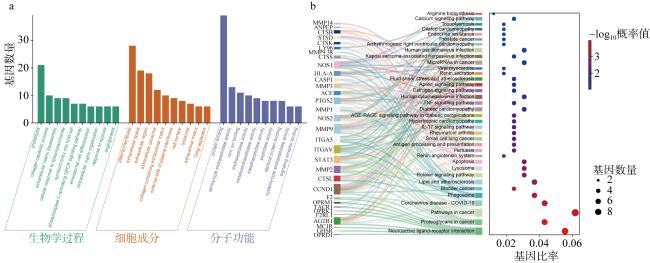
图2 海参抗骨关节炎活性肽氨基酸序列特征和靶点分析a. 骨关节炎疾病库基因靶点与潜在海参肽对应基因靶点交集Veen图; b. 24条肽特征氨基酸数量; c. 24条肽特征氨基酸占比; d. 潜在抑制骨关节炎海参活性肽的蛋白互作网络图; e. PPI中前十的基因靶点 Fig. 2 Amino acid sequence characteristics and target analysis of anti-osteoarthritis peptides from sea cucumber. (a) Veen diagram showing the intersection between OA-related gene targets and potential sea cucumber peptide targets; (b) characteristic amino acid composition of 24 peptides; (c) characteristic amino acid counts of 24 peptides; (d) protein-protein interaction (PPI) network of potential anti-OA sea cucumber peptides; (e) top ten gene targets in the PPI network |
表2 24条海参肽分子对接结合能Tab. 2 Molecular docking binding energies of 24 sea cucumber peptides |
| 肽序列 | 对接结合能(kcal·mol-1) | |
|---|---|---|
| MMP9(8K5Y) | IL-17RA(5N9B) | |
| FDPVIEEYHNGF | -9.1 | -7.0 |
| KWVF | -8.4 | -6.7 |
| SPGPY | -8.1 | -6.9 |
| DDW | -7.9 | -6.3 |
| YGRL | -7.8 | -6.6 |
| LMWCP* | -7.8 | -6.4 |
| DFPDGR | -7.5 | -6.6 |
| GHSR | -7.3 | -5.8 |
| DAW | -7.2 | -6.2 |
| TW | -7.2 | -6.7 |
| GW | -7.1 | -6.3 |
| GROW | -6.7 | -7.0 |
| APGDR | -6.6 | -6.0 |
| APR | -6.6 | -6.4 |
| AW | -6.6 | -6.2 |
| LVW | -6.6 | -6.3 |
| FAYDPAAQNKF | -6.3 | -8.0 |
| GVIF | -6.3 | -6.1 |
| NDHGF | -6.3 | -6.3 |
| ERF | -6.1 | -5.8 |
| FL | -6.1 | -5.5 |
| RR | -6.1 | -5.8 |
| SCIIP* | -6.0 | -5.3 |
| NGQYYDYSGAPSIYWSYL | -5.7 | -6.4 |
| GR | -5.6 | -5.0 |
| EF | -5.5 | -5.5 |
注: *表示该肽为已报道对骨关节炎有缓解作用的海洋生物肽, 在此为阳性肽作为筛选参考。 |
表3 化学反应描述符值Tab. 3 Chemical reactivity descriptor values |
| 参数 | KWVF | FDPVIEEYHNGF | SPGPY | LMWCP | SCIIP |
|---|---|---|---|---|---|
| EHOMO/(kJ·mol-1) | -0.00567 | -0.06577 | -0.00885 | -0.02392 | -0.03917 |
| ELUMO/(kJ·mol-1) | -0.199 | -0.2097 | -0.22609 | -0.20165 | -0.21981 |
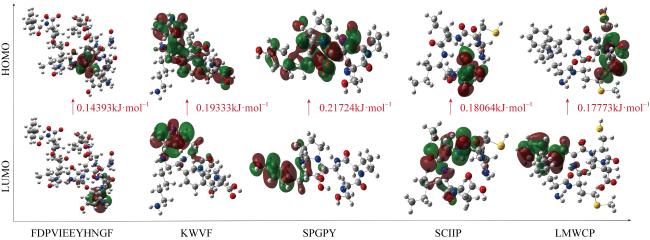
| ΔEHOMO-LUMO | 0.19333 | 0.14393 | 0.21724 | 0.17773 | 0.18064 |
| I=-EHOMO | 0.00567 | 0.06577 | 0.00885 | 0.02392 | 0.03917 |
| A=-ELUMO | 0.199 | 0.2097 | 0.22609 | 0.20165 | 0.21981 |
| χ=(I+A)/2 | 0.102335 | 0.137735 | 0.11747 | 0.112785 | 0.12949 |
| μ=-(I+A)/2 | -0.102335 | -0.137735 | -0.11747 | -0.112785 | -0.12949 |
| η= (I-A)/2 | 0.096665 | 0.071965 | -0.10862 | -0.088865 | -0.09032 |
| S= 1/η | 10.34500595 | 13.89564372 | -9.20640766 | -11.25302425 | -11.07174491 |
| ω= μ2/2η | 0.05416879 | 0.131806644 | -0.063520534 | -0.071571801 | -0.092823628 |
注: EHOMO, 最高占据分子轨道能量EHOMO能隙 (∆E); ELUMO, 最高占据分子轨道能量EHOMO能隙; ΔEHOMO-LUMO, 最高和最低占据分子轨道能差; χ, 绝对电负性; μ, 化学势; η, 绝对硬度; S, 化学软度; ω, 全局亲电性。 |
附表1 105条肽的反应活性、抗炎活性、毒性及过敏性预测结果Appendix Tab. 1 Predicted reactivity, anti-inflammatory activity, toxicity and allergenicity of 105 peptides |
| 肽序列 | 活性预测 | 抗炎活性预测 | 毒性预测 | 过敏性预测 |
|---|---|---|---|---|
| MVSLVPCGF | 0.732412 | 阴性 (0.09) | 无毒性 | 低过敏性 |
| QGMCLDVEGSDGSGNVGIY | 0.522422 | 阴性 (0.11) | 无毒性 | 低过敏性 |
| SGGSYCNGQWVDVHT | 0.515439 | 阴性 (0.15) | 无毒性 | 低过敏性 |
| AYHNTMRF | 0.809287 | 阴性 (0.18) | 无毒性 | 低过敏性 |
| CANEDCTF | 0.668399 | 阴性 (0.23) | 无毒性 | 低过敏性 |
| FTNDQF | 0.561741 | 阴性 (0.26) | 无毒性 | 低过敏性 |
| SGDLLVW | 0.687684 | 阴性 (0.26) | 毒性 | 高过敏性 |
| AVLCPPAK | 0.575315 | 阴性 (0.3) | 毒性 | 低过敏性 |
| TW | 0.814447 | —————————— | 无毒性 | 低过敏性 |
| GVIF | 0.679797 | 阳性 (0.91) | 无毒性 | 低过敏性 |
| INHCIF | 0.814483 | 阴性 (0.32) | 无毒性 | 低过敏性 |
| VF | 0.815398 | —————————— | 无毒性 | 低过敏性 |
| TW | 0.814447 | —————————— | 无毒性 | 低过敏性 |
| GVIF | 0.679797 | 阳性 (0.91) | 无毒性 | 低过敏性 |
| LVPCG | 0.555399 | 阳性 (0.97) | 无毒性 | 低过敏性 |
| NDHGF | 0.73823 | 阳性 (0.97) | 无毒性 | 低过敏性 |
| LYG | 0.571687 | 阳性 (0.98) | 无毒性 | 低过敏性 |
| SPGPY | 0.834009 | 阳性(0.69) | 无毒性 | 低过敏性 |
| IF | 0.949173 | —————————— | 无毒性 | 低过敏性 |
| IG | 0.501816 | —————————— | 无毒性 | 低过敏性 |
| LF | 0.986934 | —————————— | 无毒性 | 低过敏性 |
| LL | 0.618551 | —————————— | 无毒性 | 低过敏性 |
| LR | 0.569984 | —————————— | 无毒性 | 低过敏性 |
| MR | 0.849148 | —————————— | 无毒性 | 低过敏性 |
| IF | 0.949173 | —————————— | 无毒性 | 低过敏性 |
| TRGCMWQ | 0.562712 | 阴性 (0.33) | 无毒性 | 低过敏性 |
| SLADPAGR | 0.560956 | 阴性 (0.34) | 无毒性 | 低过敏性 |
| LYPEIPSSQRWRL | 0.73744 | 阴性 (0.36) | 无毒性 | 低过敏性 |
| LPGDNDQIPF | 0.642526 | 阴性 (0.39) | 无毒性 | 低过敏性 |
| FAYDPSAQNKF | 0.666363 | 阴性 (0.41) | 无毒性 | 低过敏性 |
| DDFPNF | 0.927081 | 阴性 (0.42) | 无毒性 | 低过敏性 |
| PHMAKDKRFEEICTKMRL | 0.767377 | 阴性 (0.47) | 无毒性 | 低过敏性 |
| CSCYPDWMAIPGSGCY | 0.929117 | 阳性 (0.54) | 毒性 | 高过敏性 |
| FTSGGMARDFPDGRGIWHNDKKNFL | 0.57984 | 阳性 (0.51) | 无毒性 | 低过敏性 |
| STW | 0.608876 | 阳性 (0.52) | 无毒性 | 低过敏性 |
| SNTAPGCAPF | 0.746688 | 阳性 (0.52) | 无毒性 | 低过敏性 |
| CSF | 0.938752 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| FGS | 0.848001 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| GAW | 0.95923 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| GPELF | 0.799842 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| NGQYYDYSGAPSIYWSYL | 0.592428 | 阳性 (0.56) | 无毒性 | 低过敏性 |
| AAAHAEGDIGGMIALQPALNGGGINHCIF | 0.530885 | 阳性 (0.56) | 无毒性 | 低过敏性 |
| GAY | 0.546287 | 阳性 (0.58) | 无毒性 | 低过敏性 |
| QNW | 0.769836 | 阳性 (0.58) | 无毒性 | 低过敏性 |
| MGGT | 0.589573 | 阳性 (0.59) | 无毒性 | 低过敏性 |
| LVW | 0.679093 | 阳性 (0.6) | 无毒性 | 低过敏性 |
| NGGG | 0.630468 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| GMALSPHVCR | 0.630588 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| PDQRF | 0.818055 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| WTN | 0.511211 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| LGR | 0.620526 | 阳性 (0.63) | 无毒性 | 低过敏性 |
| APGDR | 0.611329 | 阳性 (0.63) | 无毒性 | 低过敏性 |
| MQAIKCVVVGDGAVGKTCL | 0.596152 | 阳性 (0.64) | 无毒性 | 低过敏性 |
| TGF | 0.837013 | 阳性 (0.64) | 无毒性 | 低过敏性 |
| APR | 0.649879 | 阳性 (0.65) | 无毒性 | 低过敏性 |
| FSPGPYG | 0.903072 | 阳性 (0.65) | 无毒性 | 低过敏性 |
| MSGR | 0.598963 | 阳性 (0.66) | 无毒性 | 低过敏性 |
| PEF | 0.723457 | 阳性 (0.68) | 无毒性 | 低过敏性 |
| LPGDNDQIPF | 0.642526 | 阴性 (0.39) | 无毒性 | 低过敏性 |
| FAYDPSAQNKF | 0.666363 | 阴性 (0.41) | 无毒性 | 低过敏性 |
| DDFPNF | 0.927081 | 阴性 (0.42) | 无毒性 | 低过敏性 |
| PHMAKDKRFEEICTKMRL | 0.767377 | 阴性 (0.47) | 无毒性 | 低过敏性 |
| CSCYPDWMAIPGSGCY | 0.929117 | 阳性 (0.54) | 毒性 | 高过敏性 |
| FTSGGMARDFPDGRGIWHNDKKNFL | 0.57984 | 阳性 (0.51) | 无毒性 | 低过敏性 |
| STW | 0.608876 | 阳性 (0.52) | 无毒性 | 低过敏性 |
| SNTAPGCAPF | 0.746688 | 阳性 (0.52) | 无毒性 | 低过敏性 |
| CSF | 0.938752 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| FGS | 0.848001 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| GAW | 0.95923 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| GPELF | 0.799842 | 阳性 (0.55) | 无毒性 | 低过敏性 |
| NGQYYDYSGAPSIYWSYL | 0.592428 | 阳性 (0.56) | 无毒性 | 低过敏性 |
| AAAHAEGDIGGMIALQPALNGGGINHCIF | 0.530885 | 阳性 (0.56) | 无毒性 | 低过敏性 |
| GAY | 0.546287 | 阳性 (0.58) | 无毒性 | 低过敏性 |
| QNW | 0.769836 | 阳性 (0.58) | 无毒性 | 低过敏性 |
| MGGT | 0.589573 | 阳性 (0.59) | 无毒性 | 低过敏性 |
| LVW | 0.679093 | 阳性 (0.6) | 无毒性 | 低过敏性 |
| NGGG | 0.630468 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| GMALSPHVCR | 0.630588 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| PDQRF | 0.818055 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| WTN | 0.511211 | 阳性 (0.62) | 无毒性 | 低过敏性 |
| LGR | 0.620526 | 阳性 (0.63) | 无毒性 | 低过敏性 |
| APGDR | 0.611329 | 阳性 (0.63) | 无毒性 | 低过敏性 |
| MQAIKCVVVGDGAVGKTCL | 0.596152 | 阳性 (0.64) | 无毒性 | 低过敏性 |
| TGF | 0.837013 | 阳性 (0.64) | 无毒性 | 低过敏性 |
| APR | 0.649879 | 阳性 (0.65) | 无毒性 | 低过敏性 |
| FSPGPYG | 0.903072 | 阳性 (0.65) | 无毒性 | 低过敏性 |
| MSGR | 0.598963 | 阳性 (0.66) | 无毒性 | 低过敏性 |
| PEF | 0.723457 | 阳性 (0.68) | 无毒性 | 低过敏性 |
| PNF | 0.937503 | 阳性 (0.7) | 无毒性 | 低过敏性 |
| DEAIRAVLCPPAKNKSKRSCQ | 0.556741 | 阳性 (0.71) | 无毒性 | 低过敏性 |
| SCEDNLDQW | 0.551231 | 阳性 (0.72) | 无毒性 | 低过敏性 |
| YGRL | 0.758053 | 阳性 (0.72) | 无毒性 | 低过敏性 |
| DAW | 0.81178 | 阳性 (0.73) | 无毒性 | 低过敏性 |
| STYDNDNDVYSINCASHSSYGRGAWWYKSCL | 0.894838 | 阳性 (0.73) | 无毒性 | 低过敏性 |
| DQMW | 0.873162 | 阳性 (0.74) | 无毒性 | 低过敏性 |
| FDPVIEEYHNGF | 0.613992 | 阳性 (0.74) | 无毒性 | 低过敏性 |
| CNGQW | 0.83143 | 阳性 (0.75) | 无毒性 | 低过敏性 |
| LQPAL | 0.573567 | 阳性 (0.75) | 无毒性 | 低过敏性 |
| SDQQIIMCGDGTIRNEARNYC | 0.771888 | 阳性 (0.76) | 无毒性 | 低过敏性 |
| CNGDY | 0.565136 | 阳性 (0.77) | 无毒性 | 低过敏性 |
| KWVF | 0.877281 | 阳性 (0.78) | 无毒性 | 低过敏性 |
| TSF | 0.503464 | 阳性 (0.78) | 无毒性 | 低过敏性 |
| FAYDPAAQNKF | 0.665849 | 阳性 (0.79) | 无毒性 | 低过敏性 |
| CGNDPNAHPDGY | 0.545128 | 阳性 (0.82) | 无毒性 | 低过敏性 |
| FKCKL | 0.73601 | 阳性 (0.82) | 无毒性 | 低过敏性 |
| VWNPQIIKCTRSNTAPGCAP | 0.570157 | 阳性 (0.83) | 无毒性 | 低过敏性 |
| DFPDGR | 0.836716 | 阳性 (0.85) | 无毒性 | 低过敏性 |
| HNGF | 0.825278 | 阳性 (0.85) | 无毒性 | 低过敏性 |
| CSCYPDWMAIPGSGCYKYVDTPKT | 0.744716 | 阳性 (0.86) | 无毒性 | 低过敏性 |
| DQW | 0.7393 | 阳性 (0.87) | 无毒性 | 低过敏性 |
| GCMW | 0.990831 | 阳性 (0.88) | 无毒性 | 低过敏性 |
| GIW | 0.954702 | 阳性 (0.88) | 无毒性 | 低过敏性 |
| ERF | 0.623648 | 阳性 (0.89) | 无毒性 | 低过敏性 |
| CDDLR | 0.538337 | 阳性 (0.9) | 无毒性 | 低过敏性 |
| DSW | 0.733247 | 阳性 (0.9) | 无毒性 | 低过敏性 |
| GGG | 0.871626 | 阳性 (0.9) | 无毒性 | 低过敏性 |
| DCY | 0.64835 | 阳性 (0.94) | 无毒性 | 低过敏性 |
| DDW | 0.7639 | 阳性 (0.94) | 无毒性 | 低过敏性 |
| AF | 0.973259 | —————————— | 无毒性 | 低过敏性 |
| AW | 0.9669 | —————————— | 无毒性 | 低过敏性 |
| CF | 0.99641 | —————————— | 无毒性 | 低过敏性 |
| DF | 0.942412 | —————————— | 无毒性 | 低过敏性 |
| EF | 0.598976 | —————————— | 无毒性 | 低过敏性 |
| FL | 0.989567 | —————————— | 无毒性 | 低过敏性 |
| GF | 0.994712 | —————————— | 无毒性 | 低过敏性 |
| GG | 0.88736 | —————————— | 无毒性 | 低过敏性 |
| GH | 0.530477 | —————————— | 无毒性 | 低过敏性 |
| GL | 0.808777 | —————————— | 无毒性 | 低过敏性 |
| GR | 0.766288 | —————————— | 无毒性 | 低过敏性 |
| GW | 0.993164 | —————————— | 无毒性 | 低过敏性 |
| HF | 0.950971 | —————————— | 无毒性 | 低过敏性 |
| IC | 0.663199 | —————————— | 无毒性 | 低过敏性 |
| NF | 0.941145 | —————————— | 无毒性 | 低过敏性 |
| RL | 0.626352 | —————————— | 无毒性 | 低过敏性 |
| RR | 0.565498 | —————————— | 无毒性 | 低过敏性 |
| SW | 0.93391 | —————————— | 无毒性 | 低过敏性 |
| TF | 0.826678 | —————————— | 无毒性 | 低过敏性 |
| [1] |
孔德新, 陈雪红, 谢靖, 等, 2015. 狭鳕鱼皮胶原蛋白肽对大鼠关节炎模型中氧化应激和炎症因子的影响[J]. 安徽农业科学, 43(26): 132-134.
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}