
Journal of Tropical Oceanography >
Denitrifying microbial activity and community characteristics in coastal lagoons of South China
Received date: 2025-02-25
Revised date: 2025-04-03
Online published: 2025-04-08
Supported by
National Natural Science Foundation of China(42476144)
General Program of Guangdong Basic and Applied Basic Research Foundation(2023A1515011747)
General Program of Guangdong Basic and Applied Basic Research Foundation(2025A1515010828)
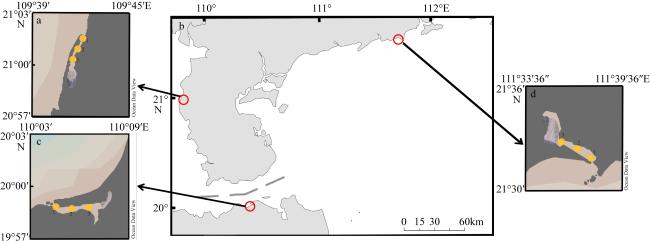
The objective of this study was to investigate the activity of denitrifying microorganisms and the factors influencing their community composition in the coastal lagoons of South China, with the aim of elucidating key mechanisms governing the nitrogen cycle in lagoon ecosystems. Three representative lagoons in South China were selected for this research. By measuring the physicochemical parameters of water and sediment, combined with denitrification rates and metagenomic sequencing data, we analyzed the community composition of three denitrifying functional genes (nirK, nirS, nosZ) and their correlations with environmental factors. The results indicated that the denitrification rate in South China lagoons was significantly lower than marine sediment averages, primarily due to low nutrient concentration, high water transparency, and limited water exchange capacity. Notably, the abundance and diversity of nirK genes within the three denitrification functional gene communities were significantly lower than those of nirS and nosZ, suggesting that nirK was more susceptible to environmental variations. Significant differences were observed in the community composition of nirK, nirS and nosZ among different lagoons. At the order level, the dominant species were Nitrosopumilales, Oceanospirillales and Flavobacteriales, respectively, with the main community composition closely resembling that of denitrifying microorganisms in the marine environment. Water transparency, pH and $\text{NH}_{4}^{+}$ concentration were identified as the key environmental factors regulating the composition of the three denitrification functional gene communities. Specifically, the nirS community was more influenced by sediment physicochemical parameters, while the nirK community was more affected by water physicochemical parameters. Additionally, the nirS and nosZ communities were similarly impacted by the same environmental factors, indicating a stronger synergistic effect in the denitrification process. This study provides valuable data to enhance our understanding of denitrification functions and microbial communities in lagoon ecosystems, thereby laying a foundation for their protection and management.
RONG Qing , XIAN Hanbiao , DONG Xuhui , YE Fei , WU Jiapeng , WANG Yu , LI Jiahui , HONG Yiguo . Denitrifying microbial activity and community characteristics in coastal lagoons of South China[J]. Journal of Tropical Oceanography, 2025 , 44(6) : 108 -119 . DOI: 10.11978/2025031
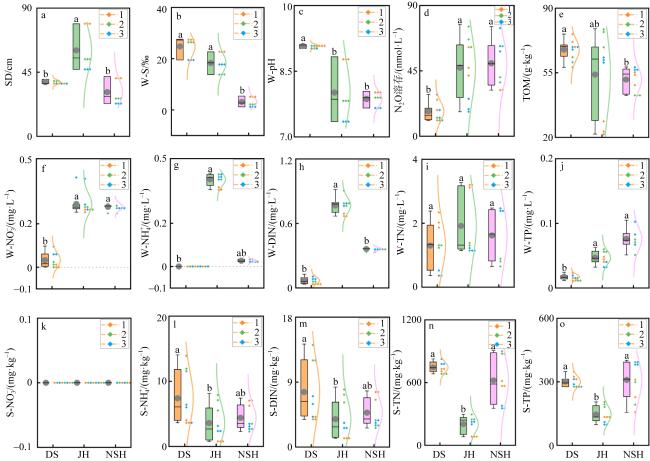
图2 三个潟湖的理化参数a. 水体透明度; b. 水体盐度; c. 水体pH; d.水体N2O溶存; e.沉积物有机质; f.水体硝酸盐氮浓度; g. 水体氨氮浓度; h.水体无机氮浓度; i.水体总氮浓度; j.水体总磷浓度; k.沉积物硝酸盐氮浓度; l.沉积物氨氮浓度; m.沉积物无机氮浓度; n.沉积物总氮浓度; o. 沉积物总磷浓度。图中灰色实心圆为平均值, 右上角图例部分的1、2、3表示潟湖对应采样点, 误差棒上方不同字母表示显著性检验结果(P<0.05), 有相同字母表示无显著差异。NSH潟湖1号位点水体遗失导致无数据 Fig. 2 Physicochemical parameters of the three lagoons. (a) Secchi depth; (b) water salinity; (c) water pH; (d) dissolution of N2O in water; (e) sediment organic matter; (f) nitrate nitrogen concentration in water; (g) ammonia nitrogen concentration in water; (h) inorganic nitrogen concentration in water; (i) total nitrogen concentration in water; (j) total phosphorus concentration in water; (k) nitrate nitrogen concentration in sediments; (l) ammonia nitrogen concentration in sediments; (m) inorganic nitrogen concentration in sediments; (n) total nitrogen concentration in sediments; (o) total phosphorus concentration in sediments |
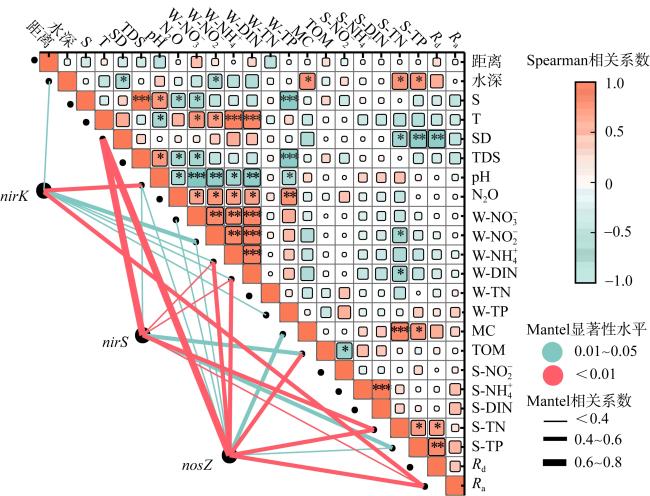
表1 各潟湖距入口距离与理化参数的Spearman相关性分析Tab. 1 Spearman correlation analysis between the distance from each lagoon to the estuary and the physical and chemical parameters |
| 潟湖 | 理化参数 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S | SD | T | pH | W-$\text{NO}_{3}^{-}$ | W-$\text{NH}_{4}^{+}$ | W-DIN | W-TN | S-$\text{NH}_{4}^{+}$ | S-$\text{NO}_{2}^{-}$ | S-DIN | S-TP | |
| DS | -1.000** | / | 1.000** | -1.000** | -0.767* | / | 0.843** | -0.949** | 0.369 | 0.847** | 0.369 | -0.525 |
| JH | -0.500 | -1.000** | 1.000** | -1.000** | -0.685* | 0.738* | 0.738* | -0.791* | 0.474 | -0.791* | 0.474 | -0.482 |
| NSH | -1.000** | -1.000** | -0.429 | -1.000** | 0.201 | -0.891* | 0.497 | 0.891* | -0.754* | -0.088 | -0.715* | 0.717* |
| 所有潟湖 | -0.369 | -0.370 | 0.158 | -0.527** | 0.407* | 0.069 | 0.350 | -0.476* | -0.070 | -0.166 | -0.035 | 0.082 |
注: *表示在0.05级别(双尾)相关性显著, **表示在0.01级别(双尾)相关性显著。 |
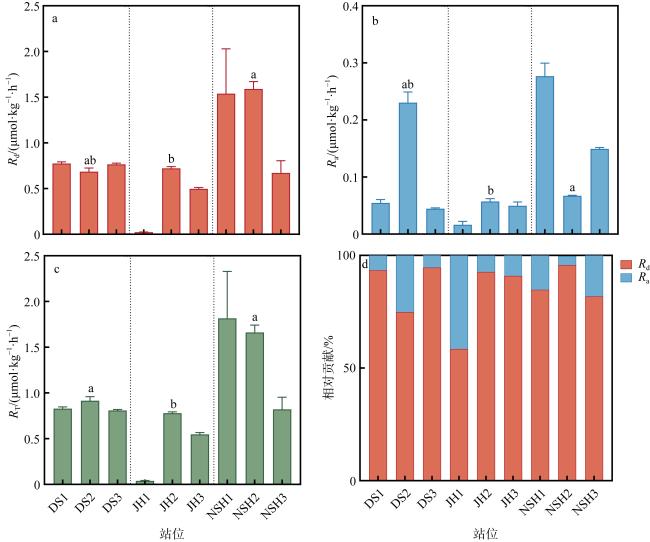
图3 三个潟湖反硝化、厌氧氨氧化和脱氮速率的比较a. 反硝化速率; b. 厌氧氨氧化速率; c. 反硝化速率和厌氧氨氧化速率为主的脱氮速率; d. 反硝化与厌氧氨氧化速率对脱氮速率的相对贡献。不同字母为显著性检验结果(P<0.05); 计算结果基于湿重进行 Fig. 3 Comparison of denitrification and anammox nitrogen removal rates in the three lagoons. (a) Denitrification rate; (b) anammox rate; (c) nitrogen removal rate dominated by denitrification and anammox rates; (d) relative contributions of denitrification and anammox rates to denitrification removal rate |
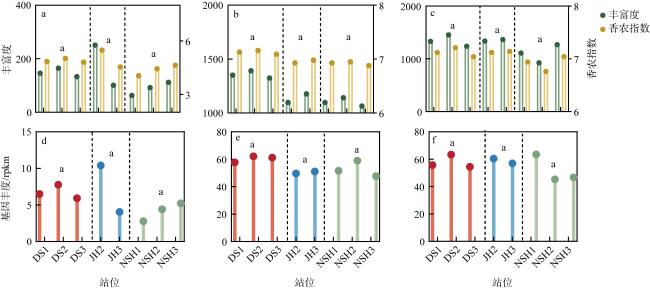
图4 三个潟湖Alpha多样性指数及基因丰度a. nirK在各位点中丰富度及香农指数; b. nirS在各位点中丰富度及香农指数; c. nosZ在各位点中丰富度及香农指数; d. 各位点nirK基因丰度; e. 各位点nirS基因丰度; f. 各位点nosZ基因丰度。同组比较时不同字母表示具有显著性差异(P<0.05)。JH潟湖1号位点的Alpha多样性指数及rpkm值异常, 图中未体现 Fig. 4 Alpha diversity indices and gene abundance in the three lagoons. (a) nirK richness and Shannon index at each site; (b) nirS richness and Shannon index at each site; (c) nosZ richness and Shannon index at each site; (d) nirK gene abundance at each site; (e) nirS gene abundance at each site; (f) nosZ gene abundance at each site |
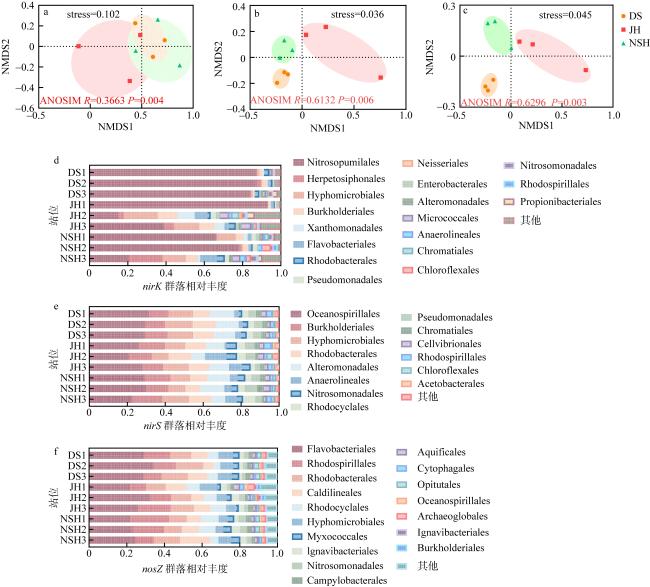
图5 三个潟湖基于OTU水平的NMDS分析及目水平群落组成a—c. nirK、nirS、nosZ基于OTU分类水平的NMDS分析; d—f. nirK、nirS、nosZ目水平群落组成 Fig. 5 NMDS analysis based on OTU level and community composition at order level in the three lagoons.(a-c) NMDS analysis of .nirK, nirS and nosZ based on OTU classification level; (d-f) NMDS analysis of.nirK, nirS and nosZ based on order classification level |
| [1] |
关凤杰, 洪义国, 吴佳鹏, 等, 2017. 发展一种小体系连续快速测定海洋水体溶解性铵盐的方法[J]. 生态科学, 36(2): 42-48.
|
| [2] |
韩磊, 胡盎, 任明磊, 等, 2023. 湖泊微生物群落及其介导的碳循环过程[J]. 生命科学, 35(12): 1613-1629.
|
| [3] |
洪义国, 张宝善, 吴佳鹏, 等, 2025. 河口近海N2O的分布特征及微生物代谢驱动机制[J]. 热带海洋学报: 1-13.
|
| [4] |
王雅君, 张司一, 欧素英, 等, 2025. 基于短时调和分析的广东惠州双月湾潟湖体系潮汐动力特征分析[J]. 热带海洋学报, 44(1): 93-107.
|
| [5] |
邢容容, 刘修锦, 邱若峰, 2019. 七里海潟湖湿地近期演变分析及生态修复研究[J]. 海洋开发与管理, 36(11): 64-68.
|
| [6] |
杨柳燕, 赵兴青, 肖琳, 等, 2006. 沉积物总氮、总磷联合测定分析方法: CN1869656[P]. 2006-11-29 (in Chinese).
|
| [7] |
杨文焕, 石大钧, 张元, 等, 2020. 高原湖泊沉积物中反硝化微生物的群落特征: 以包头南海湖为例[J]. 中国环境科学, 40(1): 431-438.
|
| [8] |
张奕璞, 于硕, 黄大吉, 等, 2022. 海南新村潟湖潮汐、潮流特征及其对营养盐的影响[J]. 海洋学研究, 40(2): 69-82.
|
| [9] |
赵锋, 许海, 詹旭, 等, 2021. 太湖春夏两季反硝化与厌氧氨氧化速率的空间差异及其影响因素[J]. 环境科学, 42(5): 2296-2302.
|
| [10] |
周茜, 彭秋萧, 张才巧, 等, 2021. 贵州草海沉积物中养分的空间分布与评价[J]. 西南大学学报(自然科学版), 43(11): 33-39.
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}