
Journal of Tropical Oceanography >
Morphological and molecular identification of five Gracilaria species (Gracilariaceae, Rhodophyta) from the Zhanjiang coastal area
Received date: 2025-02-18
Revised date: 2025-04-22
Online published: 2025-04-27
Supported by
Core Technology Research Project for Suitable Species of Modern Marine Ranch in Guangdong Province, China(2024-MRB-00-001)
Science and Technology Program of Zhanjiang, China(2022A01207)
Zhanjiang Marine Youth Talent Innovation Project, China(2023E0008)
Southern Marine Science and Engineering Guangdong Laboratory (Zhanjiang) Project, China(ZJW-2023-12)
Discipline Construction Project of Guangdong Medical University, China(4SG23009G)
Discipline Construction Project of Guangdong Medical University, China(4SG22264G)
Zhanjiang, located in the South China Sea, is rich in seaweed resources. To clarify the species diversity of the genus Gracilaria in this region, this study conducted a taxonomic investigation of intertidal macroalgae in the coastal waters of Zhanjiang using morphological, anatomical, and molecular approaches. The findings revealed the presence of five species: Gracilaria hainanensis, G. spinulosa, G. salicornia, and two as-yet undescribed species (Gracilaria sp.). The current taxonomic status of these five species was discussed, and several existing issues and controversies within the classification system were revealed. This study not only clarifies the taxonomy of Gracilaria species in the Zhanjiang coastal area, but also provides a theoretical basis for the development of red algal resources and the conservation of biodiversity in this region.
Key words: Gracilariaceae; phylogenetic analysis; rbcL; diversity; cryptic species
LI Nenghui , LYU Zhengwen , CAI Hongyan , LI Hang , ZENG Jun , HE Haiping , ZHU Yuzhen , WU Kefeng , ZHAO Wei , TAN Huaqiang , LUO Hui . Morphological and molecular identification of five Gracilaria species (Gracilariaceae, Rhodophyta) from the Zhanjiang coastal area[J]. Journal of Tropical Oceanography, 2025 , 44(6) : 74 -90 . DOI: 10.11978/2025026
附表1 各样品采集地点及其GenBank登录号Tab. S1 Sample collection locations and their GenBank accession numbers |
| 物种 | 样品编号 | 采集地点 | 登录号 | |
|---|---|---|---|---|
| rbcL | cox1 | |||
| 刺边江蓠 (Gracilaria spinulosa) | XW24051301 | 徐闻县角尾乡 | PQ799435 | PQ799449 |
| XW24051302 | 徐闻县角尾乡 | PQ799436 | PQ799450 | |
| XW24051305 | 徐闻县角尾乡 | PQ799437 | PQ799451 | |
| 海南江蓠 (Gracilaria hainanensis) | ZJ23042602 | 湛江市特呈岛 | PQ799445 | PQ799442 |
| LZ23041405 | 雷州市乌石镇 | PQ799446 | PQ799443 | |
| XW24051306 | 徐闻县角尾乡 | PP942170 | PQ799454 | |
| 缢江蓠 (Gracilaria salicornia) | LZ23041408 | 雷州市乌石镇 | PQ799447 | PQ799444 |
| LZ23041409 | 雷州市海尾村 | PQ799448 | PQ799452 | |
| Gracilaria sp.1 | LZ23053001 | 雷州市乌石镇 | PP353687 | PP353684 |
| LZ23053002 | 雷州市乌石镇 | PP353688 | PP353685 | |
| XW23061503 | 徐闻县角尾乡 | PP353689 | PP353686 | |
| Gracilaria sp.2 | LZ23041402 | 雷州市乌石镇 | PQ799431 | PQ799438 |
| LZ23041403 | 雷州市乌石镇 | PQ799432 | PQ799439 | |
| XW23061502 | 徐闻县海珠港 | PQ799433 | PQ799440 | |
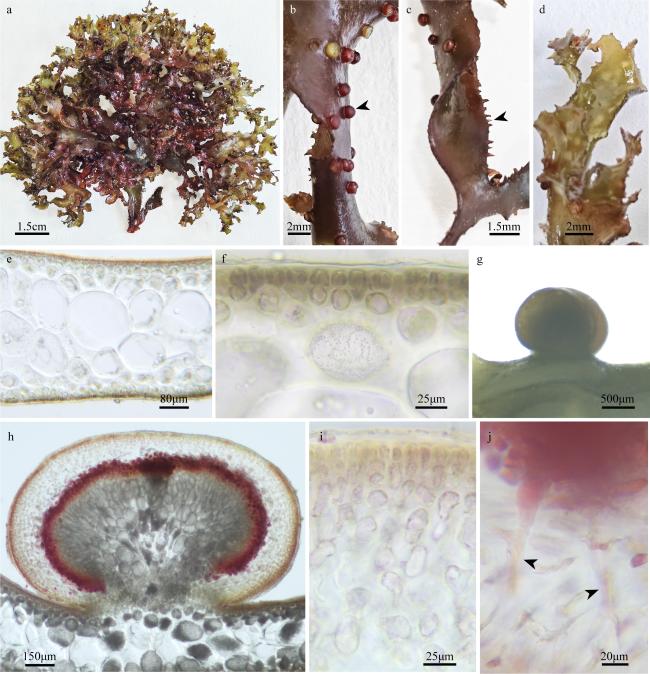
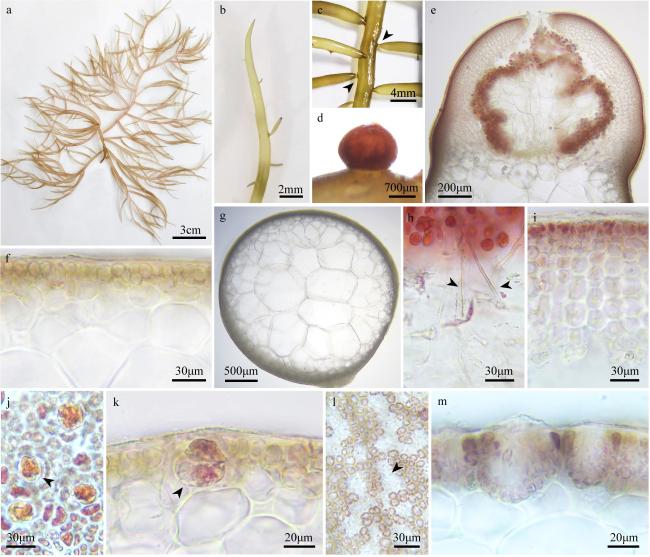
图2 刺边江蓠a. 藻体外观; b. 囊果外观(箭头); c. 叶片边缘(箭头); d. 叶片顶端; e、f. 藻体横切面观; g. 成熟囊果外观; h. 囊果切面观; i. 囊果被切面观; j. 吸收丝(箭头) Fig. 2 Gracilaria spinulosa. (a) The habit of the fresh frond; (b) the surface of the cystocarps (arrowhead); (c) the blade margin (arrowhead); (d) upper part of the blade; (e, f) the cross-section of the frond; (g) the surface of the mature cystocarp; (h) the longitudinal section of cystocarp; (i) the longitudinal section of pericarp; (j) the traversing filaments (arrowheads) |
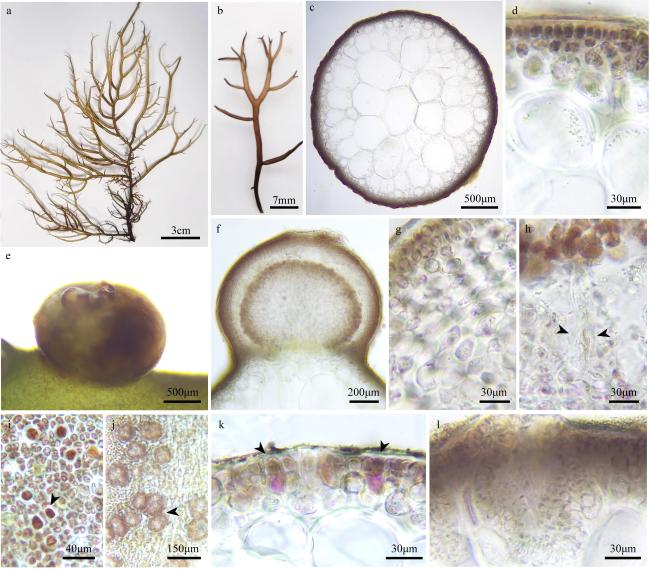
图3 海南江蓠a. 藻体外观; b. 缢缩分枝外观(箭头); c. 藻体横切面观; d. 分枝基部缢缩部位横切面观; e. 藻体部分横切面观; f. 囊果切面观; g. 囊果被切面观; h. 吸收丝(箭头); i. 精子囊窠表面观(箭头); j. 四分孢子囊表面观(箭头); k. 四分孢子囊切面观(箭头); l. 精子囊窠切面观 Fig. 3 Gracilaria hainanensis. (a) The habit of the fresh frond; (b) a branching view of constriction (arrowhead); (c) the cross-section of the frond; (d) the cross-section of the stem at the branch base; (e) the cross-section of part of the frond; (f) the longitudinal section of the cystocarp; (g) the longitudinal section of the pericarp; (h) the traversing filaments (arrowheads); (i) the surface view of spermatangia (arrowhead); (j) the surface view of tetrasporangia (arrowhead); (k) the cross-section of the tetrasporangia (arrowhead); (l) the cross-section of the spermatangia |
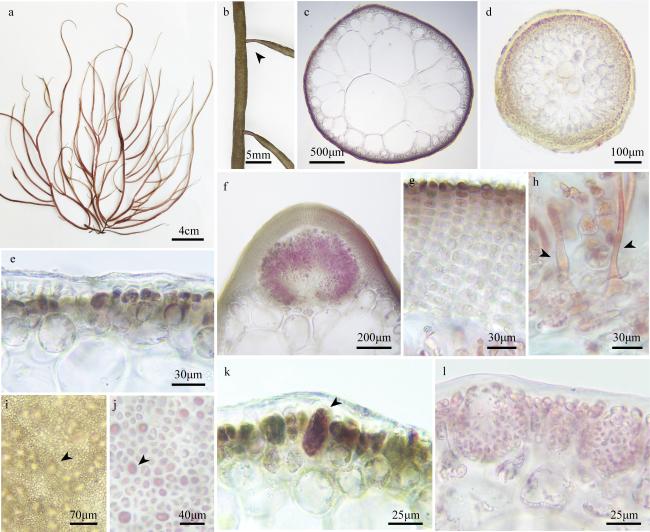
图4 缢江蓠a. 新鲜藻体表观; b、c. 分枝横切面观; d. 囊果外观; e. 吸收丝(箭头); f. 囊果切面观; g. 囊果被切面观; h. 四分孢子囊表面观(箭头); i. 四分孢子囊切面观(箭头); j. 精子囊窠表面观(箭头); k. 精子囊窠切面观 Fig. 4 Gracilaria salicornia. (a) The habit of the fresh frond; (b, c) the cross-section of the frond; (d) the surface of the cystocarp; (e) the traversing filaments (arrowheads); (f) the longitudinal section of the cystocarp; (g) the longitudinal section of the pericarp; (h) the surface view of tetrasporangia (arrowhead); (i) the cross-section of the tetrasporangia (arrowheads); (j) the surface view of spermatangia (arrowhead); (k) the cross-section of the spermatangia |
图5 Gracilaria sp.1a. 藻体外观; b. 尖锐的顶部; c. 缢缩分枝外观(箭头); d. 囊果表观; e. 囊果纵切面观; f、g. 藻体切面观; h. 吸收丝(箭头); i. 囊果被切面观; j. 四分孢子囊表面观(箭头); k. 四分孢子囊切面观(箭头); l. 精子囊窠表面观(箭头); m. 精子囊窠切面观 Fig. 5 Gracilaria sp.1. (a) The habit of the fresh frond; (b) the sharp tip; (c) a branching view of constriction (arrowheads); (d) the surface of the cystocarp; (e) the longitudinal section of the cystocarp; (f, g) the cross-section of the frond; (h) the traversing filaments (arrowheads); (i) the longitudinal section of the pericarp; (j) the surface view of tetrasporangia (arrowhead); (k) the cross-section of the tetrasporangia (arrowhead); (l) the surface view of spermatangia (arrowhead); (m) the cross-section of the spermatangia |
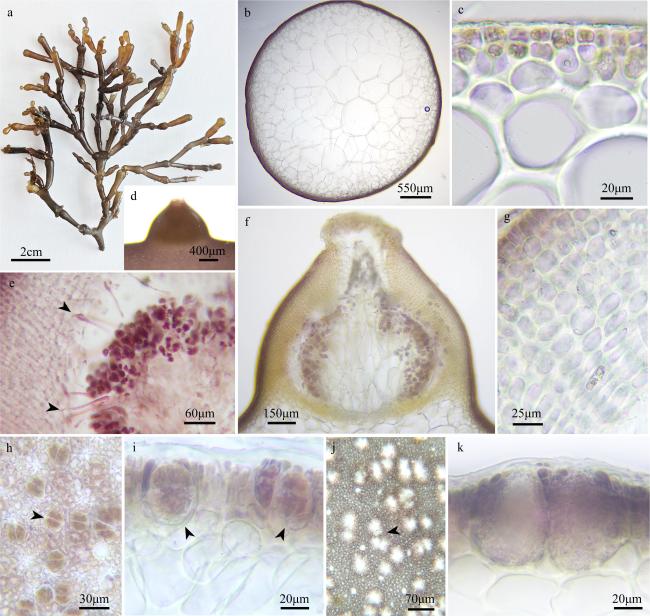
图6 Gracilaria sp.2a. 藻体外观; b. 顶部小枝; c、d. 藻体横切面观; e. 囊果外观; f. 囊果切面观; g. 囊果被切面观; h. 吸收丝(箭头); i. 四分孢子囊表面观(箭头); j. 精子囊窠表面观(箭头); k. 四分孢子囊切面观(箭头); l. 精子囊窠切面观 Fig. 6 Gracilaria sp.2. (a) The habit of the fresh frond; (b) the apical branchlet of the thallus; (c, d) the cross-section of the frond; (e) the surface of the cystocarp; (f) the longitudinal section of the cystocarp; (g) the longitudinal section of the pericarp; (h) the traversing filaments (arrowheads); (i) the surface view of tetrasporangia (arrowhead); (j) the surface view of spermatangia (arrowhead); (k) the cross-section of the tetrasporangia (arrowheads); (l) the cross-section of the spermatangia |
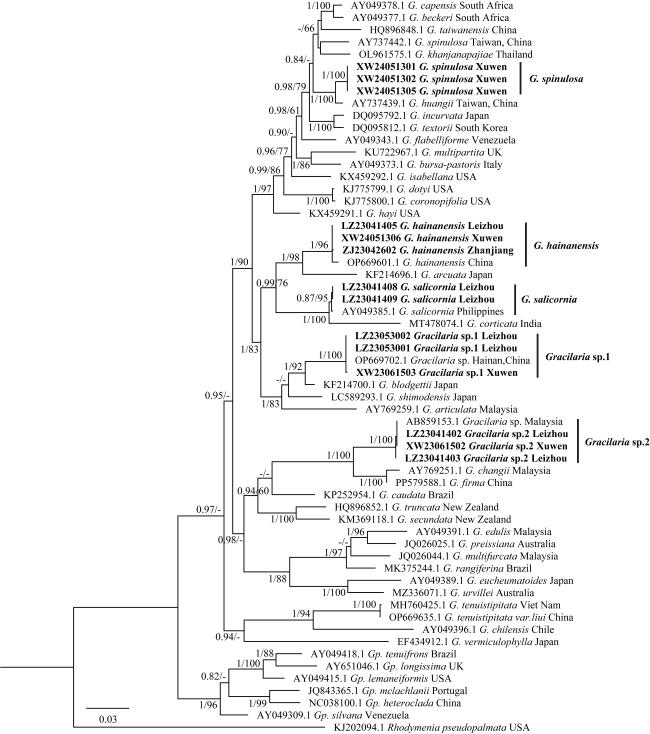
图7 基于rbcL序列构建的最大似然树黑体为本研究序列, 仅展示贝叶斯分析后验概率(PP)≥0.6/最大似然分析自展值(BP)≥60的节点数值; “-”表示低于0.6PP和60%BP以下的支持值 Fig. 7 Maximum likelihood tree based on the rbcL sequence. The bold text in the figure represents the sequences from this study. Only nodes with Bayesian posterior probabilities (PP)≥0.6 or maximum likelihood bootstrap percentages (BP)≥60 are shown; A '-' indicates support values below 0.6 PP and 60% BP |
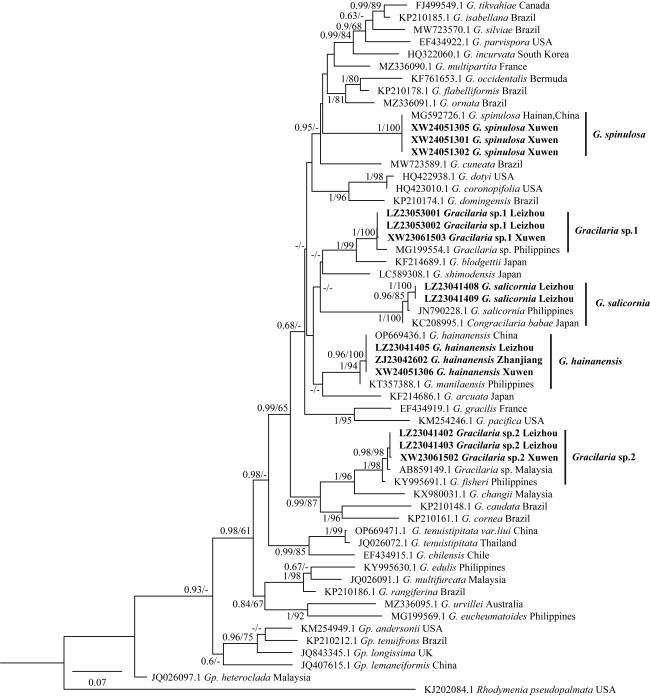
图8 基于cox1序列构建的最大似然树黑体为本研究序列, 仅显示贝叶斯分析后验概率(PP)≥0.6/最大似然分析自展值(BP)≥60的节点数值; “-”表示低于0.6PP和60%BP以下的支持值 Fig. 8 Maximum likelihood tree based on the cox1 sequence. The bold text in the figure represents the sequences from this study. Only nodes with Bayesian posterior probabilities (PP)≥0.6 or maximum likelihood bootstrap percentages (BP)≥60 are shown; A '-' indicates support values below 0.6 PP and 60% BP |
表1 样品与刺边江蓠及G. huangii的形态特征比较Tab. 1 Comparison of morphological characteristics between the samples and G. spinulosa, and G. huangii |
| 特征 | 样品 | 刺边江蓠 | 刺边江蓠(台湾) | G. huangii | |
|---|---|---|---|---|---|
| 高/cm | 7~12 | 6~11.5 | 2.8~9 | 4~8 | |
| 宽/cm | 0.5~2.5 | 0.5~1.2 | 0.2~0.7 | 0.4~1.5 | |
| 体厚/μm | 288~562 | 415~448 | 110~160 (~405) | 250~500 | |
| 固着器 | 不规则盘状, 具短柄 | 不规则盘状, 具短柄 | 盘状, 偶有短柄 | 盘状, 上有短柄 | |
| 分枝模式 | 分枝密集; 不规则的二叉式分枝 | 不规则的二叉式分枝 | 疏松至密集的不规则二叉式分枝 | 分枝疏松; 3~5次不规则二叉式分枝 | |
| 髓部 | 3~5层; 75~216μm | 4~5层; 直径达132μm | 2~3层; 75~90(~120)μm×90~120 (~195)μm | 1~3层; 75~160μm×150~170μm | |
| 表层 | 1~2层; 6.6~12.7μm×5.4~10.8μm | 2层; 7~10μm×3~7μm | 1~2层; 5~8μm×5~6μm | 1~2层; 5~6μm | |
| 内皮层 | 1~2层; 15~42μm | 20~40μm | 1~2层; 30~65μm | 1~2层; 12~20μm | |
| 囊果大小 | 近球形; 0.7~1.1mm×0.9~1.6mm | 近球形; 1.2mm×1.6mm | 半球形; 1.1~1.4mm | 半球形; 1.2~2.2mm | |
| 囊果被 | 12~15层; 厚219~345μm | 12~14层; 厚332~415μm | 未观察 | 未观察 | |
| 吸收丝 | 明显 | 明显 | 明显 | 稀少 | |
| 精子囊窠 | 未观察 | “T”型; 33~40μm×26~43μm | “T”型 | “T”型 | |
| 四分孢子囊 | 未观察 | 未观察 | 散布藻体皮层 | 散布藻体皮层 | |
| 参考文献 | 本文 | 夏邦美(1999) | Lin (2006a) | Lin et al (2006b) | |
表2 样品与海南江蓠、硬江蓠及G. manilaensis的形态特征比较Tab. 2 Comparison of morphological characteristics between the samples and G. hainanensis, G. firma, and G. manilaensis |
| 特征 | 样品 | 海南江蓠 | G. manilaensis | 硬江蓠 |
|---|---|---|---|---|
| 高/cm | 15~46 | 15~35 | 60或更高 | 10~20 |
| 宽/mm | 1.8~3 | 2~3 | 1.5 | 1~3 |
| 分枝模式 | 不规则的互生或偏生 | 不规则互生或偏生 | 互生, 有时偏生或不规则 | 多为互生, 偶有偏生 |
| 分枝长度 | 35cm | 可达45cm | 分枝众多且较长 | 未观察 |
| 分枝缢缩 | 基部强烈缢缩 | 强烈缢缩成细柄 | 基部处明显缢缩 | 基部渐细成极细的柄 |
| 髓部 | 450~1017μm | 500~550 (~950)μm | 最大可达570μm | 230~450μm |
| 表层 | 1~3层; 8.5~15.7μm×6.3~13.5μm | 1~3层; 7~9μm×5~7μm | 1~2层; 7.2~12.8μm×8.8~13.6μm | 2~3层; 6.6~13.2μm×3.3~6.6μm |
| 内皮层 | 1~2层; 18~43μm | 23~33μm | 未观察 | 3~5层; 26~83μm |
| 囊果大小 | 锥型; 608μm×831μm | 锥形; 高414~435μm | 球形; 可达1000μm | 圆锥或半球形; 580~630μm |
| 囊果被 | 10~14层; 厚101~207μm | 9~11层; 厚90~120μm | 未观察 | 8~10层; 厚83~95μm |
| 吸收丝 | 发达 | 明显 | 存在 | 不存在 |
| 精子囊窠 | “V”型; 46~85μm×31~73μm | “V”型; 37μm×39μm | “V”型; 71μm×50μm | “V”型; 66~116μm×33~66μm |
| 四分孢子囊 | 22~42μm×15~24μm | 37~39μm×21~26μm | 35~40μm×24~27μm | 50~66μm×26~43μm |
| 参考文献 | 本文 | 张峻甫 等(1976) | Yamamoto et al (1994) | 张峻甫 等(1976) |
表3 样品与繖房江蓠及G. fisheri的形态特征比较Tab. 3 Comparison of morphological characteristics between the samples and G. coronopifolia, and G. fisheri |
| 特征 | 样品 | 繖房江蓠 | G. fisheri |
|---|---|---|---|
| 高/cm | 10~20 | 7~8 | 15~30 |
| 宽/mm | 1.3~2.8 | 1~1.5 | 0.5~2 |
| 分枝模式 | 不规则互生或叉分 | 不规则的二叉式分枝 | 3~4次互生, 偶尔侧生 |
| 分枝缢缩 | 通常不缢缩, 偶有略缩 | 不缢缩 | 轻微缢缩, 或不缢缩 |
| 髓部/μm | 208~750 | 183~365 | 240~560 |
| 表层 | 1~2层; 8.2~13.5μm×5.5~11.8μm | 1~2层; 7~10μm×4~10μm | 3~4层 |
| 内皮层 | 1~2层; 18~47μm | 2~3层, 较小 | 未观察 |
| 囊果大小 | 锥型或半球形; 588~917μm×710~1124μm | 半球形或近球形; 830~930μm×1328~1411μm | 圆锥形或半球形; 700μm |
| 囊果被 | 7~13层; 厚101~181μm | 6~10层; 厚116~162μm | 未观察 |
| 囊果被细胞形态 | 扁圆或呈星型 | 圆或扁圆 | 扁压, 内容物呈星型 |
| 果孢子囊/μm | 22~30 | 23.1~30 | 20~25 |
| 吸收丝 | 明显 | 明显 | 稀少 |
| 精子囊窠 | “P”型; 50~132μm×41~127μm | “V”型; 46~50μm×33~43μm | “P”型; 40~63μm×43~50μm |
| 四分孢子囊 | 31~40μm×13~23μm | 未观察 | 12.5~15μm×26~30μm |
| 参考文献 | 本文 | 张峻甫 等(1976) | Xia et al (1987); Abbott et al (1991) |
| [1] |
丁兰平, 黄冰心, 谢艳齐, 2011. 中国大型海藻的研究现状及其存在的问题[J]. 生物多样性, 19(6): 798-804.
|
| [2] |
丁兰平, 黄冰心, 王宏伟, 2015. 中国海洋红藻门新分类系统[J]. 广西科学, 22(2): 164-188.
|
| [3] |
丁兰平, 杨楠, 刘金梅, 等, 2020. 海洋经济红藻江蓠科的分类研究现状及存在的问题[J]. 应用海洋学学报, 39(1): 144-151.
|
| [4] |
李永梅, 刘瑞, 杨楠, 等, 2018. 海南省4种江蓠属(红藻门)海藻的形态分类学研究[J]. 热带海洋学报, 37(4): 29-37.
|
| [5] |
李能辉, 黄庆, 李航, 等, 2024. 基于形态和分子数据对湛江海域4个江蓠物种(红藻门)的分类学研究[J]. 热带海洋学报, 43(2): 34-47.
|
| [6] |
刘金梅, 黄冰心, 马鑫, 等, 2021. 沙菜属(红藻门杉藻目)的分类学问题探讨[J]. 海洋科学, 45(8): 109-117.
|
| [7] |
王波, 游伟伟, 王项愿, 等, 2010. 不同饵料对杂色鲍幼鲍生长及存活的影响[J]. 厦门大学学报(自然科学版), 49(2): 286-292.
|
| [8] |
夏邦美, 1999. 中国海藻志: 第二卷红藻门第五册杉藻目[M]. 北京: 科学出版社 (in Chinese).
|
| [9] |
谢恩义, 申玉春, 叶宁, 等, 2009. 流沙湾的底栖大型海藻调查[J]. 广东海洋大学学报, 29(4): 30-35.
|
| [10] |
薛姣, 李婷, 苏家齐, 等, 2024. 广东10个海岛潮间带大型经济海藻调查[J]. 南方水产科学, 20(4): 98-106.
|
| [11] |
于杰, 戴晓玲, 章增林, 等, 2017. 细基江蓠和真江蓠野生种群生长海域水质营养盐特征及富营养化评价[J]. 南方农业学报, 48(8): 1511-1517.
|
| [12] |
张才学, 周伟男, 孙省利, 等, 2020. 硇洲岛大型海藻群落的季节演替[J]. 热带海洋学报, 39(1): 74-84.
|
| [13] |
张峻甫, 夏邦美, 1962. 中国江蓠属植物地理学的初步研究[J]. 海洋与湖沼, 4(3-4): 189-198.
|
| [14] |
张峻甫, 夏邦美, 1976. 中国江蓠属海藻的分类研究[J]. 海洋科学集刊, 11: 91-166 (in Chinese).
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}