
Journal of Tropical Oceanography >
Bathyconchoecia nanshaensis sp. nov. (Myodocopa, Halocyprididae), a new species of ostracod from the southern South China Sea*
Online published: 2021-04-29
Supported by
National Natural Science Foundation of China(41976112)
National Natural Science Foundation of China(31971432)
National Natural Science Foundation of China(41576125)
Fund for strategic biological resources, Chinese Academy of Sciences(KFJ-BRP-017-48)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0401)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0405)
Science and Technology Basic Resources Investigation Program of China(2017FY201404)
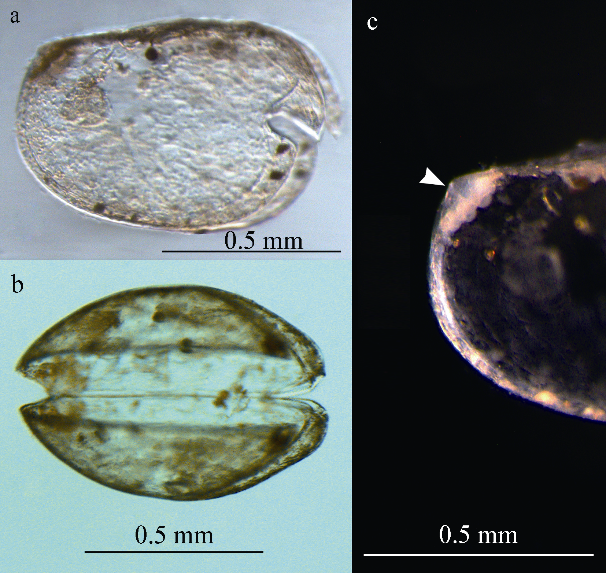
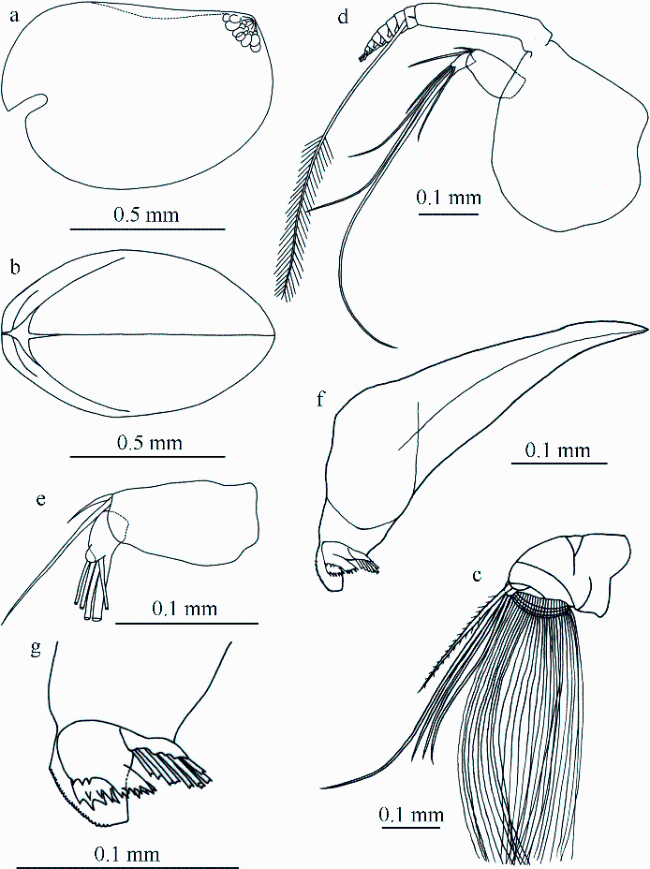
A new species of halocyprid ostracod, Bathyconchoecia nanshaensis sp. nov., from the Nansha Trough (the North-West Borneo Trough), southern South China Sea is illustrated and described in detail. The species is a small one that closely resembles Bathyconchoecia paulula Deevey, 1968 and Bathyconchoecia pacifica Chavtur, 1977 in terms of its carapace length (CL; <1.0 mm) and relative height (~70% CL). However, it can be easily distinguished from these species by the locations of its asymmetrical glands’ opening, by the structure of the coxal toothed edge of the mandible, and by the setation of the sixth limb.
Key words: Bathyconchoeciinae; systematics; zooplankton; Nansha Trough; Pacific Ocean
YIN Jianqiang , LI Kaizhi , TAN Yehui . Bathyconchoecia nanshaensis sp. nov. (Myodocopa, Halocyprididae), a new species of ostracod from the southern South China Sea*[J]. Journal of Tropical Oceanography, 2022 , 41(2) : 193 -197 . DOI: 10.11978/2021042
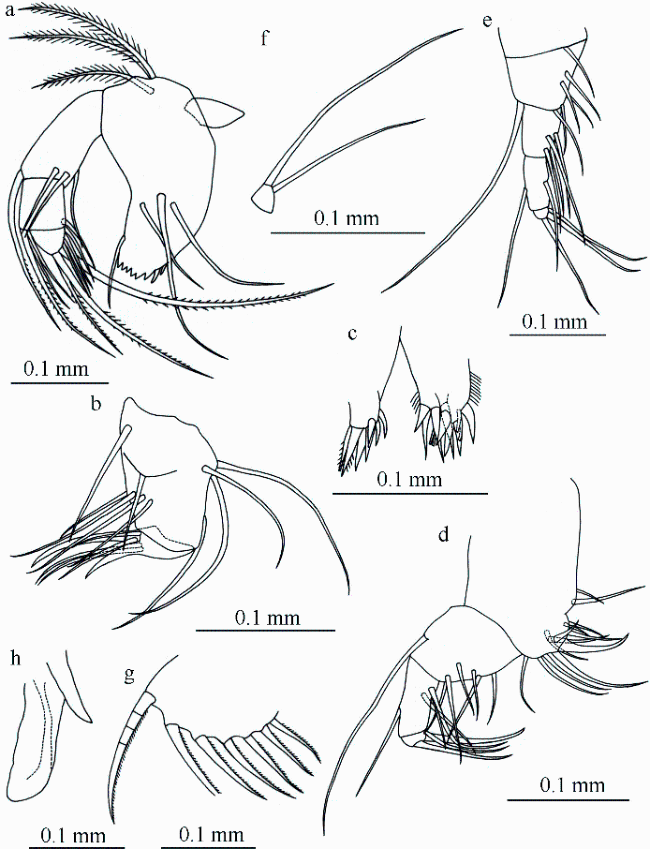
Fig. 3 Bathyconchoecia nanshaensis sp. nov., A-1 male, holotype.(a) Basale and endopodite of mandible; (b) maxilla (without endites shown in c); (c) precoxal endite (left) and coxal endite (right) of maxilla; (d) fifth limb; (e) sixth limb; (f) seventh limb; (g) caudal furca; (h) copulatory appendage |
Tab. 1 Comparison of the main morphological characteristics of male Bathyconchoecia paulula, Bathyconchoecia pacifica, and Bathyconchoecia nanshaensis sp. nov. |
| Characteristics | B. paulula (♂) | B. pacifica (♂) | B. nanshaensis (A-1 ♂) |
|---|---|---|---|
| Carapace | |||
| Length/mm | 0.95 | 0.90~0.97 | 0.87 |
| Height/Length/% | 70 | 70 | 69 |
| Rostrum | Slightly bent downwards | Sharply bent downwards | Slightly bent downwards |
| Asymmetric glands | Opening at posterodorsal corner | Opening at posterodorsal corner | Opening on posterior margin near to posterodorsal corner |
| Mandible | |||
| Coxal toothed edge | Several cutting teeth | Several cutting teeth | Smooth |
| Basal toothed edge | Six triangular cutting teeth | Five triangular cutting teeth | Six triangular cutting teeth |
| Sixth limb | |||
| Basal ventral setae | Three setae | Three setae | Five setae |
| Basal distodorsal seta (residual exopodite) | Seta barely reaches to the end of this limb | Seta does not reach to the end of this limb | Seta extends beyond the end of this limb |
| Terminal setae | Dorsal seta is 2.8 times the length of the ventral seta* | Dorsal seta is approximately three times the length of the ventral seta* | Dorsal, central, and ventral setae are subequal in length |
| Caudal furca | There is an unpaired seta | There is no unpaired seta | Unpaired seta is missing |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}