
Journal of Tropical Oceanography >
Effects of salinity on the growth and transcriptome of euryhaline Synechococcus sp. K1*
Received date: 2021-04-06
Revised date: 2021-05-01
Online published: 2021-05-03
Supported by
Nature Science Foundation of China(41906131)
Key Deployment Program of Center for Ocean Mega-Science, Chinese Academy of Sciences(COMS2020Q09)
Guangdong Basic and Applied Basic Research Foundation(2019A1515011340)
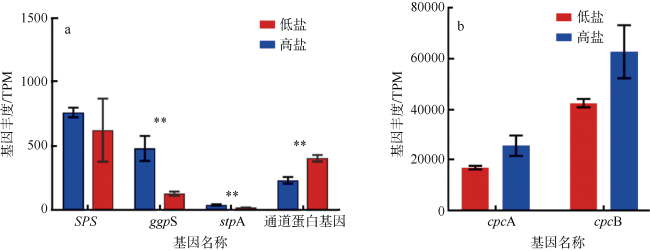
Synechococcus, which is one of the representative groups of picocyanobacteria, is widely distributed in global oceans and estuaries. It has high phenotypic (pigment) and genetic diversity. According to the level of ability to deal with variation in salinity, Synechococcus can be divided into the strictly marine type and euryhaline type. In this study, we compared the growth of Synechococcus sp. K1 (isolated from the Pearl River estuary) and Synechococcus sp. YX02-1 (isolated from the South China Sea oligotrophic water) under a series of salinity gradient conditions. We also conducted phylogenetic analysis of K1 and YX02-1 based on rpoC1 gene sequences. We found that euryhaline Synechococcus sp. K1 could grow in all salinity levels, while strictly marine Synechococcus sp. YX02-1 was unable to survive below 13‰, which is consistent with the distribution characteristics in the estuary. Transcriptome analysis showed that the expression of genes (ggpS, SPS, stpA) that synthesize osmotic pressure regulator molecules in euryhaline Synechococcus were significantly down-regulated, while the gene expression of the membrane channel protein glzT was significantly up-regulated under low-salinity condition. This suggests that the low-salt tolerance mechanism of euryhaline Synechococcus mainly includes reducing the synthesis of small molecules related to intracellular osmotic pressure and increasing membrane channel proteins to improve the efflux of small molecules. In addition, salinity affected the photosynthesis and metabolism levels of euryhaline Synechococcus. The expression of photosynthesis energy-saving genes (ATPF0B, ATPF0A, ATPF1D, ATPF1A) and pigment protein genes (cpcA, cpcB) were significantly down-regulated under low-salinity condition, and the expression of inorganic nitrogen utilization-related genes were up-regulated. Under low-salinity condition, the demand for small molecules related to osmotic pressure is reduced, which can make more carbon sources to support growth. Meanwhile, the absorption of inorganic nitrogen is enhanced. These two reasons may be responsible for the higher growth of Synechococcus under low-salinity condition.
Key words: Euryhaline Synechococcus; transcriptome analysis; salinity
LIAO Ying , XIA Xiaomin . Effects of salinity on the growth and transcriptome of euryhaline Synechococcus sp. K1*[J]. Journal of Tropical Oceanography, 2022 , 41(2) : 159 -169 . DOI: 10.11978/2021040
表1 无硅f/2培养基配方Tab. 1 The medium formula of f/2 without silicon |
| 成分 | 质量浓度/(g·L-1) | |
|---|---|---|
| 常量元素 | NaNO3 | 7.5×10 -2 |
| NaH2PO4H2O | 5.0×10 -3 | |
| 微量金属 | FeCl3·6H2O Na2EDTA2·H2O CuSO4·5H2O Na2MoO4·2H2O ZnSO4·7H2O CoCl2·6H2O MnCl4·4H2O | 3.15×10 -3 4.36×10 -3 9.8×10 -3 6.3×10 -3 2.2×10 -2 1.0×10 -2 1.8×10 -1 |
| 维生素 | Thiamine HCl (vit. B1) Biotin (vit. H) Cyanocobalamin (vit. B12) | 2.0×10 -4 1.0×10 -3 1.0×10 -3 |
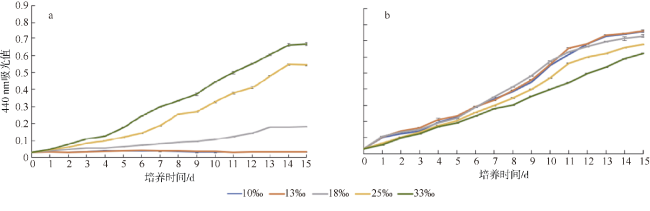
图1 聚球藻YX02-1 (a)和K1 (b)在不同盐度(10‰、13‰、18‰、25‰、33‰)下的生长曲线Fig. 1 The growth curve of Synechococcus sp. of YX02-1 (a) and K1 (b) at different salinity (10‰, 13‰, 18‰, 25‰, 33‰) |
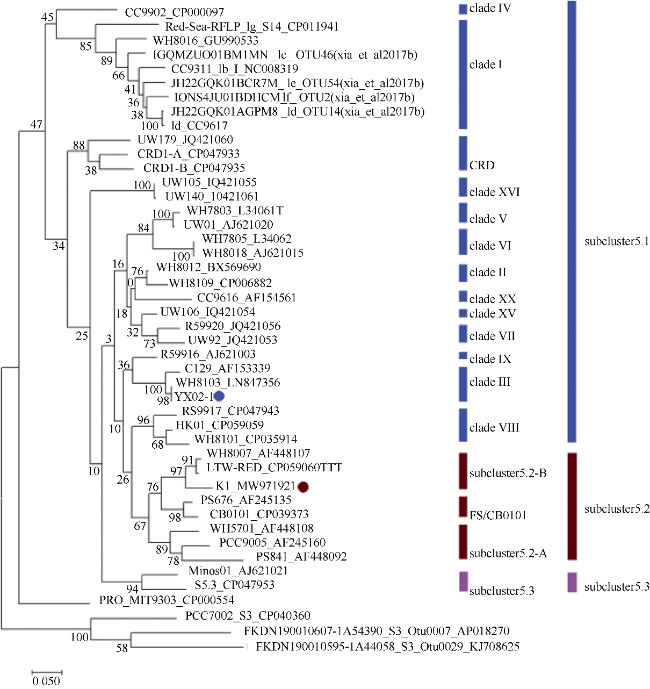
图4 聚球藻系统发育进化树主要分为3个分支: subcluster 5.1、subcluster 5.2和subcluster 5.3。聚球藻K1由红色圆圈标记, 聚球藻YX02-1由蓝色圆圈标记 Fig. 4 The phylogenetic tree of Synechococcus being mainly divided into three groups: subcluster 5.1, subcluster 5.2 and subcluster 5.3. K1 is marked by red dot, while YX02-1 is marked by blue dot |
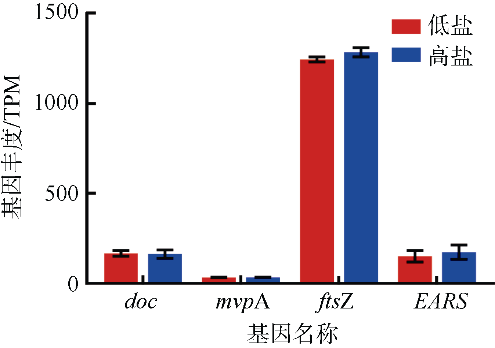
图5 热图显示低盐条件下聚球藻K1显著上调(a)或显著下调(b)的基因及丰度图中H1、H2、H3表示高盐组; L1、L2、L3表示低盐组。Sqrt(TPM)表示经过开平方转换的TPM值。图中左侧为基因名称@该基因在KEGG通路中的K号; 右侧字母为基因功能代码, J: 翻译、核糖体结构和生物发生; K: 转录; L: 复制, 重组和修复; D: 细胞周期, 细胞分裂, 染色体分配; V: 防御机制; T: 信号转导机制; M: 细胞壁/膜/包膜生物发生; N: 细胞运动; U: 细胞内运输, 分泌和囊泡转运; O: 翻译后修饰, 蛋白质周转, 分子伴侣; C: 能量产生和转化; G: 碳水化合物转运和代谢; E: 氨基酸转运和代谢; F: 核苷酸转运和代谢; H: 辅酶转运和代谢; I: 脂质转运和代谢; P: 无机离子转运和代谢; Q: 次级代谢产物生物合成, 转运和分解代谢; S: 未知功能 Fig. 5 The heatmap showing that the gene abundance (TPM) of Synechococcus sp. K1 was significantly up-regulated (a) or down-regulated (b) at low salinity |
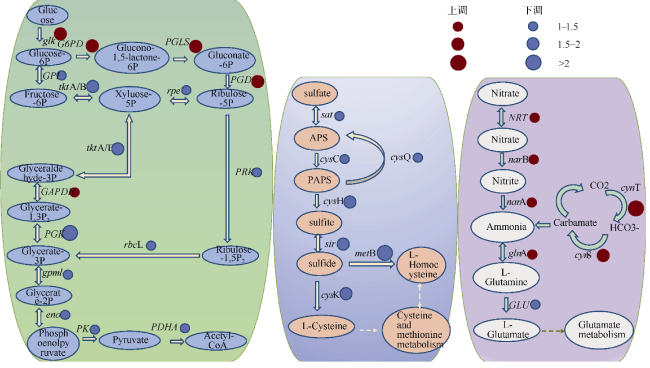
图7 聚球藻K1碳代谢(a)、硫代谢(b)、氮代谢(c)途径对盐度变化的响应红色圆圈代表基因上调, 蓝色圆圈代表基因下调。圆圈大小表示调整幅度 Fig. 7 The responses of carbon metabolism (a), sulfur metabolism (b) and nitrogen metabolism (c) of Synechococcus sp. K1 to salinity changes. The red dot represents the genes of up-regulation, and the blue dot represents the genes of down-regulation |
*感谢厦门大学郑强博士提供的实验藻株, 感谢编辑老师和审稿专家提供的宝贵意见和帮助。
| [1] |
路荣昭, 王淑芝, 关志英, 等, 1991. 聚球藻(Synechococcus leopoliensis 625)藻胆体-类囊体膜光谱特性和光能传递的研究[J]. 水生生物学报, 15(4): 368-371.
|
| [2] |
马英, 焦念志, 2004. 聚球藻(Synechococcus)分子生态学研究进展[J]. 自然科学进展, 14(9): 967-972.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}