
Journal of Tropical Oceanography >
Spatiotemporal variation of water area with high chlorophyll a concentration in the South China Sea based on OC-CCI data*
Received date: 2021-04-12
Revised date: 2021-07-24
Online published: 2021-07-30
Supported by
Subproject B of the Pilot Special Project of the Chinese Academy of Sciences(XDB42010201)
Basic Research Funds of the Second Institute of Oceanology, Ministry of Natural Resources of China(HYGG2002)
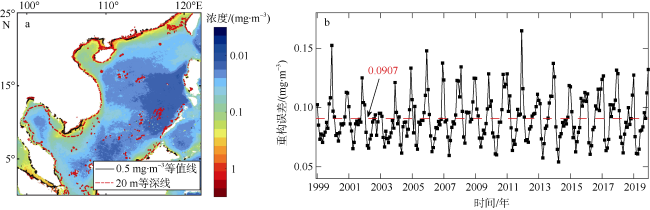
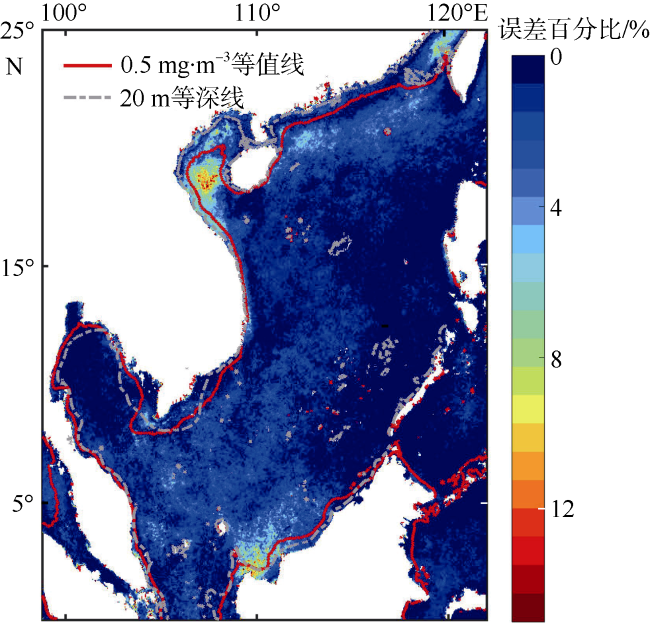
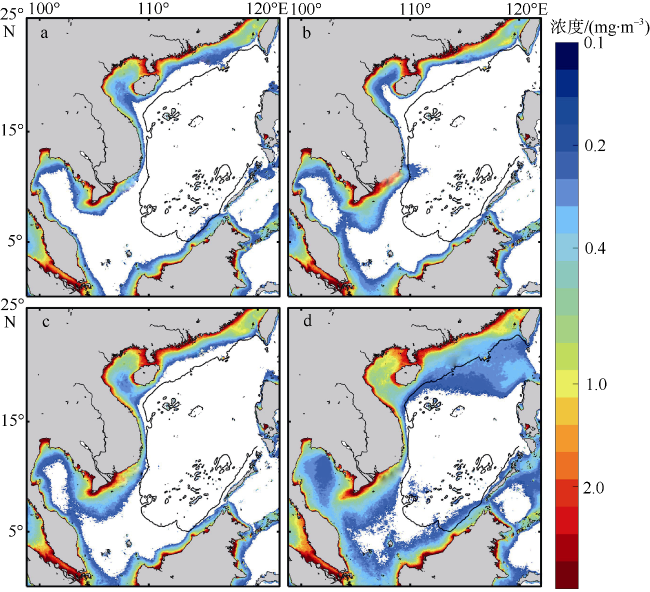
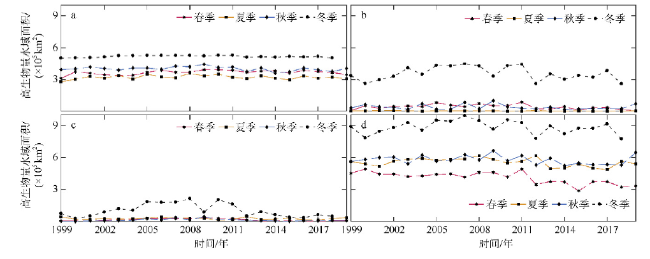
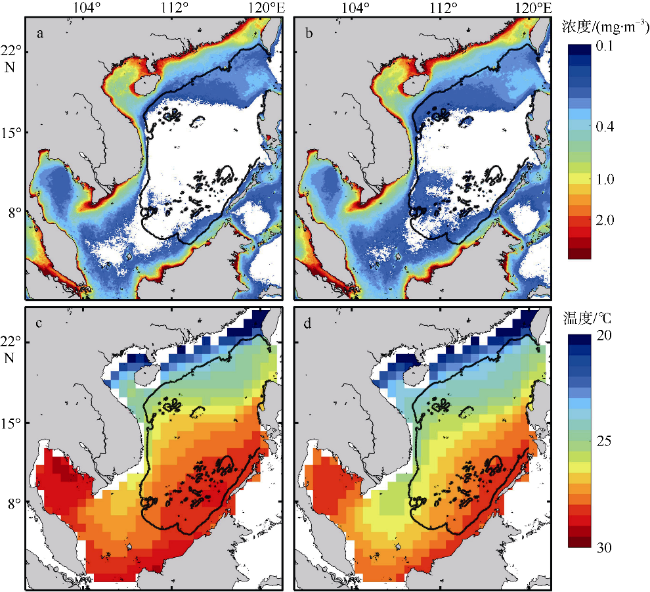
Phytoplankton are the basis of the marine ecosystem food chain, and affect the variation of CO2 flux through photosynthesis. In this study, an estimation system of phytoplankton biomass in the South China Sea was established based on an area indicator. We used the empirical orthogonal function decomposition interpolation method (DINEOF) to reconstruct the chlorophyll a concentration field in the South China Sea from long time series of remote sensing data. We studied the space-time distribution of the high biomass water area of the South China Sea and found that the changes of water area with high chlorophyll a concentration had significant seasonal characteristics. The area of waters with high chlorophyll concentration reached the maximum in winter, and the minimum in summer. Conversely, the chlorophyll concentration reached the minimum in winter, and maximum in summer. This feature may be due to the wind-driven dynamic processes that redistribute the chlorophyll concentration near the surface. Moreover, waters with high chlorophyll concentration were found near the coast year round, especially in coastal waters of China, along the coast of Vietnam, the Gulf of Thailand, and near Borneo Island. In the Sunda Shelf and the central basin east of the Mekong estuary, the area of high biomass water showed interannual variation. The East Asian monsoon modulated by the El Niño and Southern Oscillation led to different degrees of cold water invasion from the north to the southeast of the Mekong Estuary in different years, which may be the reason for the increase and decrease of local phytoplankton biomass.
LI Ao , FENG Yang , WANG Yuntao , XUE Huijie . Spatiotemporal variation of water area with high chlorophyll a concentration in the South China Sea based on OC-CCI data*[J]. Journal of Tropical Oceanography, 2022 , 41(2) : 77 -89 . DOI: 10.11978/2021048
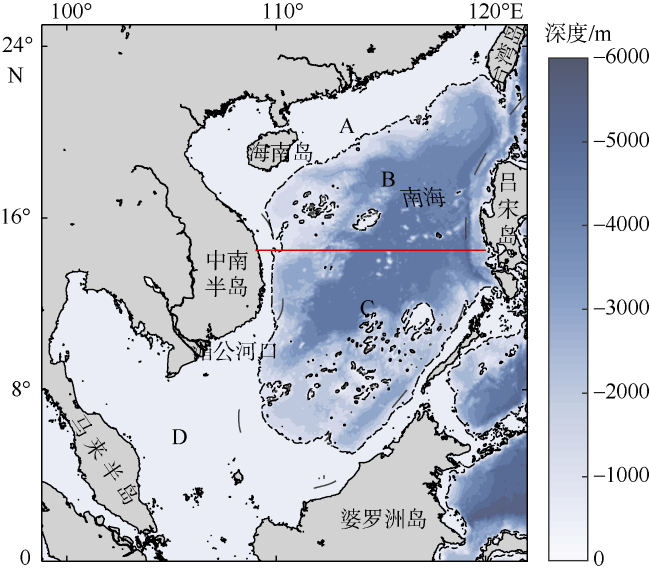
图1 南海地形分布该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392的标准地图制作。图中黑色虚线为500m等深线, 红色实线为划分南海南北的14°30′纬线。从北向南将南海划分为4个区域, 其中A区域为北部陆架区、B区域为北部海盆区、C区域为南部海盆区、D区域为南部陆架区 Fig. 1 Topographic map of South China Sea. The black dotted line shows the 500 m isobath. The solid red line (14°30′N) separates the South China Sea into north and south parts |
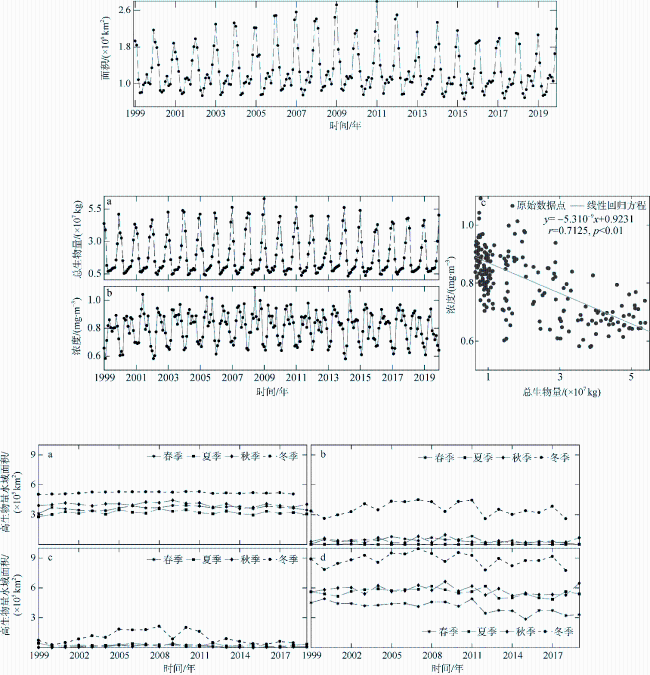
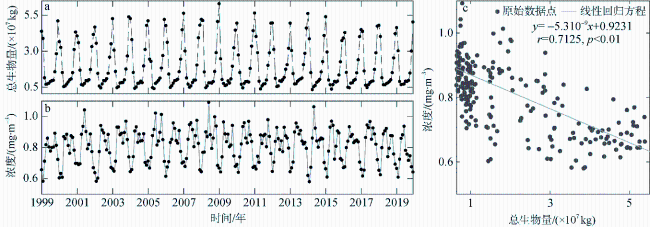
图9 南海高叶绿素a浓度水域的生物量和海表叶绿素a浓度a. 高叶绿素a浓度水域混合层生物量估算值; b. 高叶绿素a浓度水域海表叶绿素浓度; c. 高叶绿素a浓度水域混合层生物量与海表叶绿素浓度回归分析 Fig. 9 Biomass of high chlorophyll a concentration waters and chlorophyll a concentration of high chlorophyll a concentration waters in the South China Sea. (a) estimated biomass of mixed layer in high chlorophyll a concentration waters; (b) the concentration of chlorophyll a on the surface of high chlorophyll a concentration waters; (c) regression analysis between the biomass of the mixed layer and the chlorophyll a concentration at the surface with high chlorophyll a concentration |
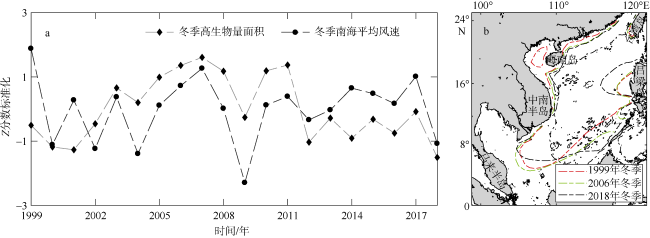
图12 南海冬季高生物量水域面积与风场的标准化时间序列(a)和不同年份海 |
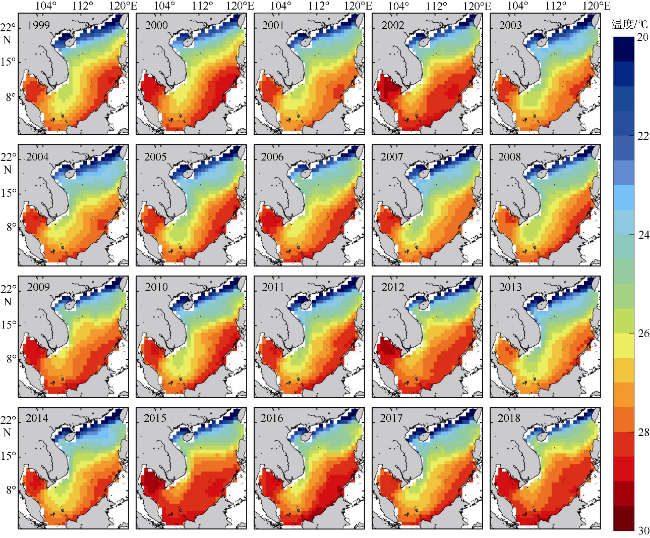
图13 南海冬季海表温度空间分布该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392的标准地图制作 Fig. 13 Spatial distribution of winter SST in the South China Sea |
*在研究过程中得到中国科学院南海海洋研究所舒婵、房巍巍、梁爱琳、胡晓悦和厦门大学电科学院李波的诸多建议和帮助, 在此表示十分感谢!感谢NASA/Ocean Color网站提供的海洋颜色气候变化倡议(OC-CCI) 5.0版本的海表月平均叶绿素a产品、欧洲气象中心提供的风场与温度数据以及日本海洋-地球科技研究所提供的混合层深度数据。
| [1] |
陈兴群, 陈其焕, 庄亮钟, 1989. 南海中部叶绿素a分布和光合作用及其与环境因子的关系[J]. 海洋学报, 11(3): 349-355.
|
| [2] |
倪晓波, 黄大吉, 2006. 海洋次表层叶绿素最大值的分布和形成机制研究[J]. 海洋科学, 30(5): 58-64, 70.
|
| [3] |
赵辉, 齐义泉, 王东晓, 等, 2005. 南海叶绿素浓度季节变化及空间分布特征研究[J]. 海洋学报, 27(4): 45-52.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}