
Journal of Tropical Oceanography >
Comparative study on photophysiology of four macroalgae from the Zhongsha Atoll, with special reference to the effects of temperature rise*
Copy editor: LIN Qiang
Received date: 2021-08-18
Revised date: 2021-10-08
Online published: 2021-10-12
Supported by
National Science & Technology Fundamental Resources Investigation Program(2018FY100104)
National Natural Science Foundation(41890853)
Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0407)
Natural Science Foundation of Guangdong Province(2022A1515011461)
Science and Technology Planning Project of Guangdong Province(2017B020217002)
Macroalgae, together with corals, provide faunal foods and habitats in clear tropical waters, maintaining the rich biological diversity in coral reefs. In this paper, we compared biochemical compositions and photosynthetic features, as well as photophysiological responses to temperature rise of two red algae (i.e., Grateloupia ellipitica and Liagora samaensis) and two green algae (i.e., Udotea flabellum and Halimeda discoidea), the dominating macroalgae species in the Zhongsha Atoll of the South China Sea. Results showed that the red algae contain less chlorophyll a and carotenoids than the green ones, but contain the unique phycoerythrin and phycocyanin. Both the light compensation point (EC) and dark respiration (Rd) of the red algae are lower than the green algae. The temperature rise enhances EC, Rd and light utilization efficiency (α), as well as the daily carbon fixation of both red and green algae; and it lowers the saturation irradiance (EK) of the former algae species but enhances the maximum photosynthetic oxygen evolution rate (Pmax) of the later species. Moreover, the temperature rise enhances the photosynthetic rate of the red algae under lower light, but reduces it under higher light, while such temperature-induced enhancement on the photosynthetic rate of the green algae occurs throughout the measured light range. Considering the inter-specific photophysiological characteristics of the four macroalgae as well as the effects of temperature, the temperature rise would benefit the photosynthesis of macroalgae in the Zhongsha Atoll and benefit the red algae better than the green algae.
LI Gang , WAN Mingyue , SHI Xiaohan , QIN Geng , MAI Guangming , HUANG Liangmin , TAN Yehui , ZOU Dinghui . Comparative study on photophysiology of four macroalgae from the Zhongsha Atoll, with special reference to the effects of temperature rise*[J]. Journal of Tropical Oceanography, 2022 , 41(3) : 101 -110 . DOI: 10.11978/2021105
表1 4种大型海藻藻体色素与蛋白含量Tab. 1 Pigments of chlorophyll a (Chl a), carotenoids (Car), phycoerythrin (PE), phycocyanin (PC), and soluble proteins (SP) contents (mg·g-1 FW) in G. ellipitica, L. samaensis, U. flabellum, and H. discoidea. Shown are the mean and standard deviation of three or four independent measurements (n=3 or 4) |
| 色素 | 蛋白含量/(mg·g-1 FW) | |||
|---|---|---|---|---|
| 厚膜藻G. ellipitica | 粉枝藻L. samaensis | 钙扇藻U. flabellum | 仙掌藻H. discoidea | |
| 叶绿素a(Chl a) | 0.03±0.006a | 0.08±0.005a | 0.36±0.068b | 0.16±0.034c |
| 类胡萝卜素(Car) | 0.004±0.001a | 0.034±0.005b | 0.26±0.056c | 0.14±0.029d |
| 藻红蛋白(PE) | 0.65±0.021a | 0.010±0.008b | — | — |
| 藻蓝蛋白(PC) | 0.004±0.003a | 0.009±0.001b | — | — |
| 可溶性蛋白(SP) | 1.12±0.087a | 1.25±0.383a | 5.17±0.533b | 1.28±0.197a |
注: 上标不同字母表示差异性显著(P<0.05) |
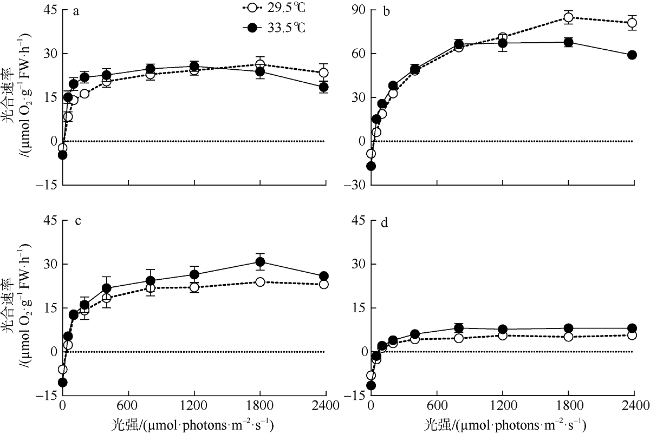
图2 原位温度和升温条件下4种大型海藻的光响应(P-E)曲线a. 厚膜藻; b. 粉枝藻; c. 钙扇藻; d. 仙掌藻 Fig. 2 Photosynthetic oxygen evolution rate (µmol O2·g-1 FW·h-1) versus irradiance (µmol·photons·m-2·s-1) (P vs. E) curves under ambient (29.5℃) and elevated temperatures (33.5℃) for (a) G. ellipitica, (b) L. samaensis, (c) U. flabellum, and (d) H. discoidea. Symbols show the averaged values of three or four independent measurements; vertical bars indicate standard deviation (n=3 or 4), often fall within the symbols |
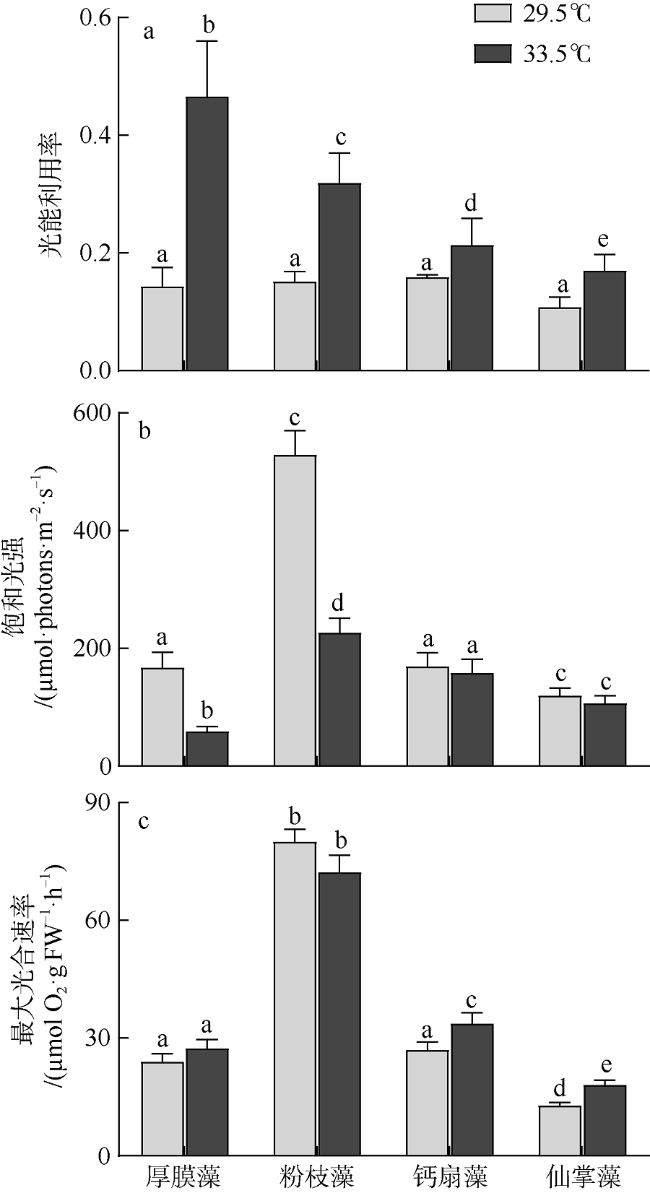
图3 原位温度(29.5℃)和升温(33.5℃)条件下4种大型海藻的光合生理参数不同字母表示差异性显著(P<0.05) Fig. 3 P vs. E curve-derived photosynthetic parameters under ambient (29.5℃) and elevated (33.5℃) temperatures for G. ellipitica, L. samaensis, U. flabellum, and H. discoidea. The letter above each bar indicates significant difference (P<0.05) |
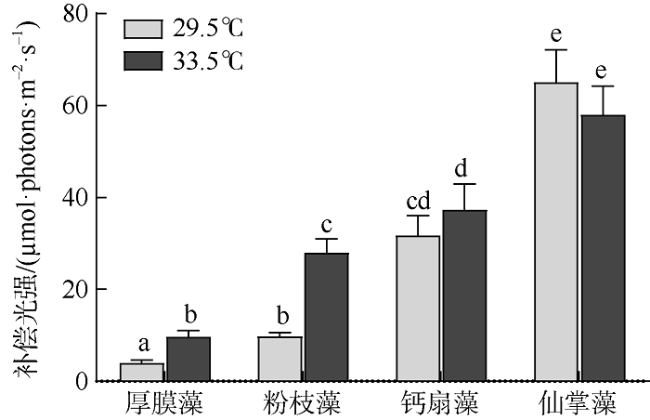
图4 原位温度(29.5℃)和升温(33.5℃)条件下4种大型海藻光补偿点变化不同字母表示差异性显著(P<0.05) Fig. 4 Light compensation point (EC, µmol·photons·m-2·s-1) under ambient (29.5℃) and elevated temperatures (33.5℃) for G. ellipitica, L. samaensis, U. flabellum and H. discoidea. Vertical bar indicates a standard deviation (n=3 or 4); the letter on top of each bar indicates the significant difference (P<0.05) |
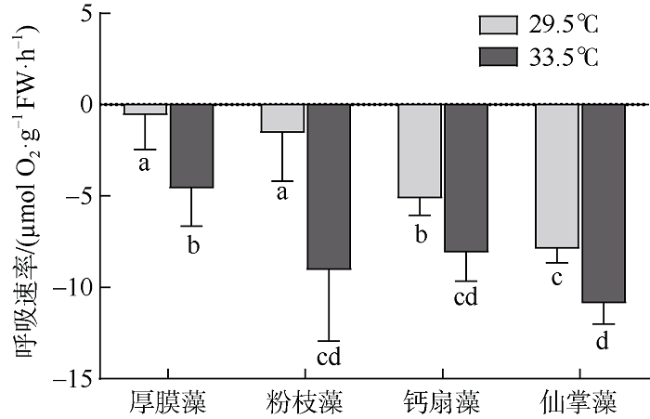
图5 原位温度(29.5℃)和升温(33.5℃)条件下4种大型海藻暗呼吸速率变化不同字母表示差异性显著(P<0.05) Fig. 5 Dark respiration (Rd; µmol O2·g-1 FW·h-1) of G. ellipitica, L. samaensis, U. flabellum, and H. discoidea under ambient (29.5℃) and elevated (33.5℃) temperatures. Vertical bar indicates standard deviation (n=3 or 4), and the letter indicates significant difference (P<0.05) |
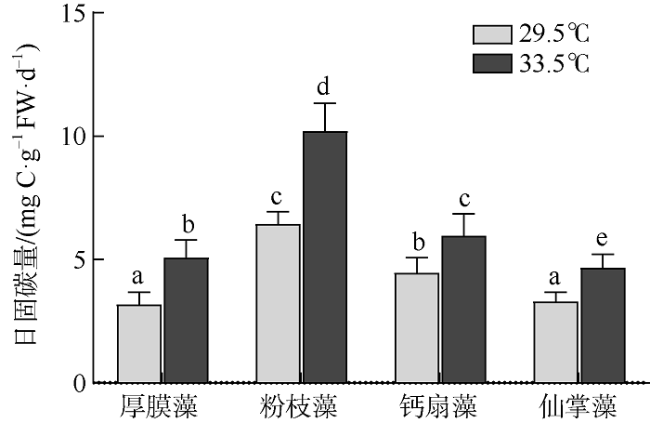
图6 原位温度和升温条件下四种大型海藻的日净光合固碳量变化不同字母表示差异性显著(P<0.05) Fig. 6 Daily carbon fixation (mg C·g-1 FW·d-1) of G. ellipitica, L. samaensis, U. flabellum, and H. discoidea under ambient (29.5℃) and elevated (33.5℃) temperatures. Vertical bar indicates standard deviation (n = 3 or 4), and the letter indicates significant difference (P<0.05) |
| [1] |
蔡逸洵, 温嘉怡, 邹定辉, 2021. 干出状态下坛紫菜叶状体表面水膜与失水对光合作用光化学特性的影响[J]. 热带海洋学报, 40(1): 82-90.
|
| [2] |
程丽巍, 邹定辉, 郑青松, 等, 2010. 光照和温度对氮饥饿及饱和营养条件下石莼(Ulva lactuca)的硝态氮吸收动力学影响[J]. 生态学杂志, 29(5): 939-944.
|
| [3] |
丁兰平, 黄冰心, 谢艳齐, 2011. 中国大型海藻的研究现状及其存在的问题[J]. 生物多样性, 19(6): 798-804.
|
| [4] |
高坤山, 2014. 藻类固碳--理论、进展与方法[M]. 北京: 科学出版社: 1-491.
|
| [5] |
黄道建, 黄小平, 岳维忠, 2005. 大型海藻体内TN和TP含量及其对近海环境修复的意义[J]. 台湾海峡, 24(3): 316-321.
|
| [6] |
李刚, 2009. 中国南海浮游植物光合固碳与阳光紫外辐射关系的研究[D]. 汕头: 汕头大学: 1-171.
|
| [7] |
龙超, 罗肇河, 韦章良, 等, 2021. 海南三亚鹿回头虫黄藻(Effrenium voratum)的形态学和系统发育学研究[J]. 热带海洋学报, 40(4): 35-43.
|
| [8] |
钱树本, 2014. 海藻学[M]. 青岛: 中国海洋大学出版社: 1-822.
|
| [9] |
沈乃澂, 2013. 南海诸岛的面积测量(续)[J]. 中国计量, (11): 62-64.
|
| [10] |
吴超元, 张京浦, 温宗存, 等, 1996. 青岛三种海产红藻的光合和呼吸特性的初步研究[J]. 海洋与湖沼, 27(2): 207-212.
|
| [11] |
杨宇峰, 2016. 近海环境生态修复与大型海藻资源利用[M]. 北京: 科学出版社: 1-364.
|
| [12] |
杨雨玲, 李伟, 陈伟洲, 等, 2013. 不同温度及二氧化碳浓度下培养的龙须菜光合生理特性对阳光紫外辐射的响应[J]. 生态学报, 33(18): 5538-5545.
|
| [13] |
章守宇, 向晨, 周曦杰, 等, 2018. 枸杞岛海藻场6种大型海藻光合荧光特性比较[J]. 应用生态学报, 29(10): 3441-3448.
|
| [14] |
周百成, 郑舜琴, 曾呈奎, 1974. 几种绿藻、褐藻和红藻的吸收光谱的比较研究[J]. 植物学报, 16(2): 146-155.
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
IPCC, 2014. Summary for policymakers[M]//IPCC. Climate Change 2013 - The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press: 1-30.
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}