
Journal of Tropical Oceanography >
Molecular cloning and expression analysis of Commd1 under salinity stress in Crassostrea hongkongensis
Received date: 2016-03-01
Request revised date: 2016-08-26
Online published: 2017-01-19
Supported by
National Natural Science Foundation of China (31572661, 31202021, 31272658)
Science and Technology Program of Guangzhou, China (201605120657132)
Copyright
COMMDs are recently discovered in many multicellular organisms. They are involved in numerous aspects of biological processes, such as the regulation of copper homeostasis, the transport of sodium ions, the activity of the transcription factors NF-κB, and hypoxia-inducible factor (HIF-1). To investigate the role of Commd1 in response to salt stress, we first cloned Commd1 cDNA using Rapid Amplification of the cDNA Ends (RACE) technique in mollusks, Crassostrea hongkongensis (designed ChCommd1). The full-length cDNA is 841 bp containing a 5′-untranslated region (UTR) in 18 bp, a 3′-UTR in 262 bp with a poly (A) tail, and an open reading frame (ORF) in 564 bp; the ORF encodes a 187 amino-acid polypeptide with a predicted molecular mass of 21.79 kDa and an isoelectric point of 5.21. Homologous comparison and phylogenetic analysis revealed that the 187-aa-long ChCOMMD1 protein shares high sequence identity with its homologs from other species and belongs to the molluscan COMMD1 family. Quantitative real-time PCR analysis showed that ChCommd1 mRNA is broadly expressed in various tissues and during different stages of the oyster’s embryonic and larval development. Upon exposure to two stressors (high and low salinity), the expression level of ChCommd1 mRNA increased significantly. Taken together, our results indicated that ChCommd1 can function in the embryonic development and osmotic regulation, providing an important reference for further investigations on the functions of Commd1 in the euryhaline mollusks.
Key words: C. hongkongensis; Commd1; gene expression; salinity stress
WANG Fuxuan , XIAO Shu , XIANG Zhiming , YU Ziniu . Molecular cloning and expression analysis of Commd1 under salinity stress in Crassostrea hongkongensis[J]. Journal of Tropical Oceanography, 2017 , 36(1) : 48 -55 . DOI: 10.11978/2016022
Tab. 1 Sequences of designed primers used in this study表1 引物信息 |
| 名称 | 序列 (5'-3') | 用途 |
|---|---|---|
| ChCommd1-R1 | TTGCTGCCTCTCTCCTCTTCATTTG | 5'RACE |
| ChCommd1-R2 | ACATCTCTTCAGTAGTGTTGCGTAA | |
| ChCommd1-F1 | CGTGTTACAAAATATGAAAGACATT | 3'RACE |
| ChCommd1-F2 | GTGCCATAGTTGAACTCCAAATAGA | |
| ChCommd1-F3 | GAAAACATGACAAACAAAATGG | ORF扩增 |
| ChCommd1-R3 | TGAGTACGGGAATATGATACAA | |
| ChCommd1-F4 | AAGCCATTGCTTACAGAAAGTT | 荧光定量PCR |
| ChCommd1-R4 | TCAGGGTTTTCTATTTGGAGTT | |
| GAPDH- F | GGATTGGCGTGGTGGTAGAG | 内参基因 |
| GAPDH- R | GTATGATGCCCCTTTGTTGAGTC | |
| EF1α-F | CGGGATCCATGTATAGTCGGGAGA | 内参基因 |
| EF1α-R | CCCAAGCTTTCACAGAGAAATCAA |
注: F表示正向引物, R表示反向引物 |
Fig. 1 Nucleotide and amino acid sequences of ChCommd1图1 ChCommd1 全长cDNA核苷酸序列及其编码的氨基酸序列 |
Fig. 2 Multiple alignment of the putative amino acid sequence of ChCommd1 with its homologues in other organisms图2 香港牡蛎与其他物种COMMD1氨基酸序列的多重比对 |
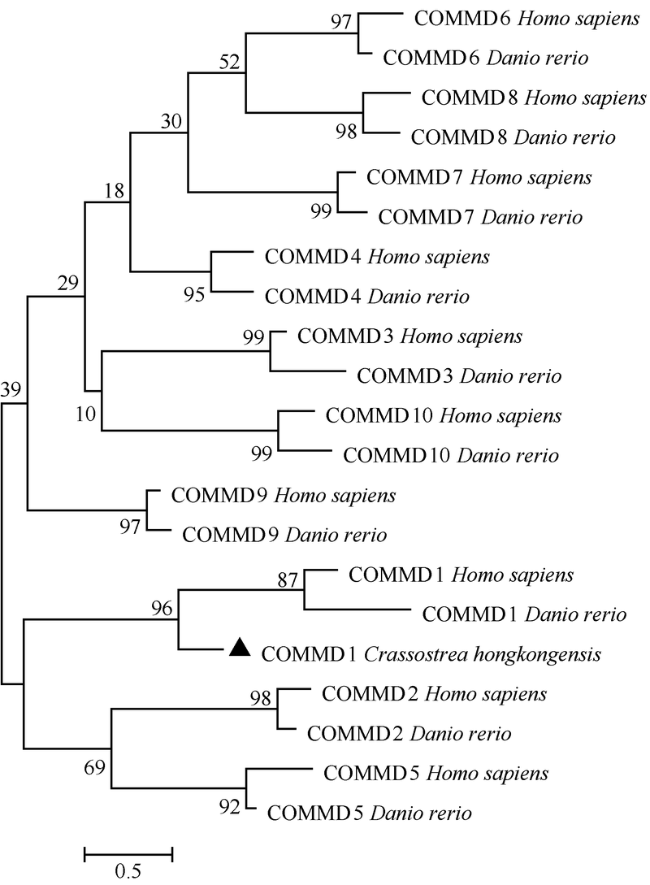
Fig. 3 Maximum-likelihood phylogenetic tree of ChCommd1 amino acid sequences图3 基于ML法构建的ChCOMMD1系统进化树 |
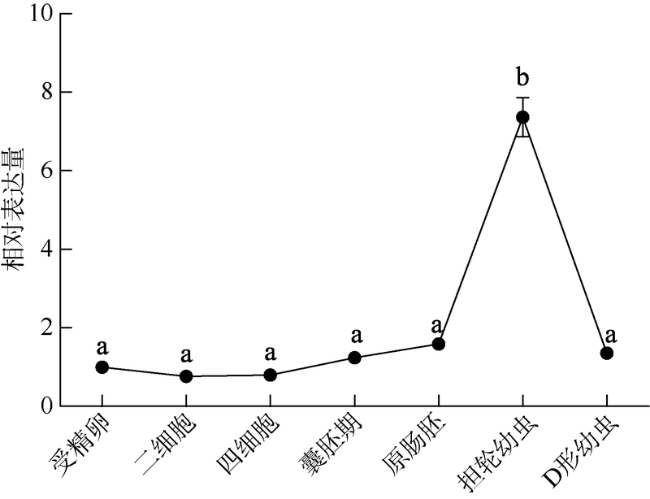
Fig. 4 Expression profiles of ChCommd1 during different stages of oyster’s embryonic and larval development图4 ChCommd1在不同胚胎发育时期的表达模式 |
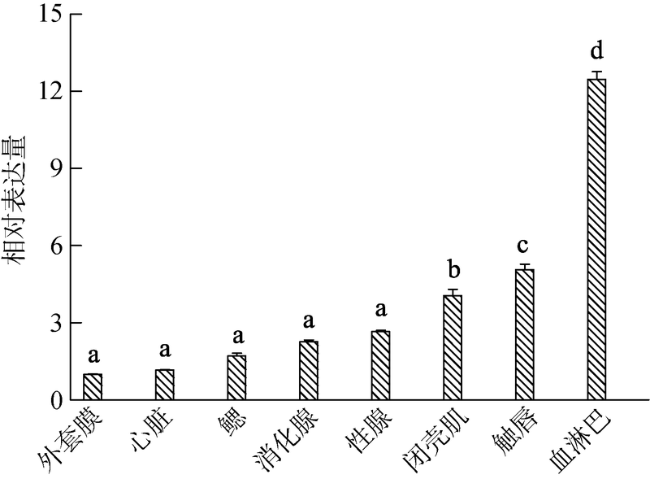
Fig. 5 Expression profiles of ChCommd1 in different adult tissues图5 ChCommd1在成贝不同组织中的表达模式 |
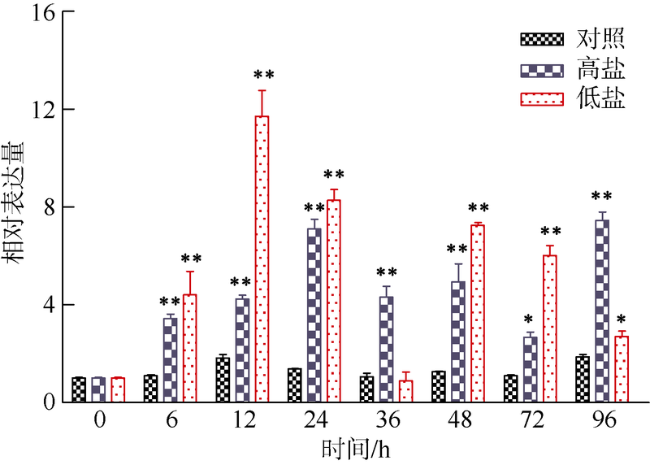
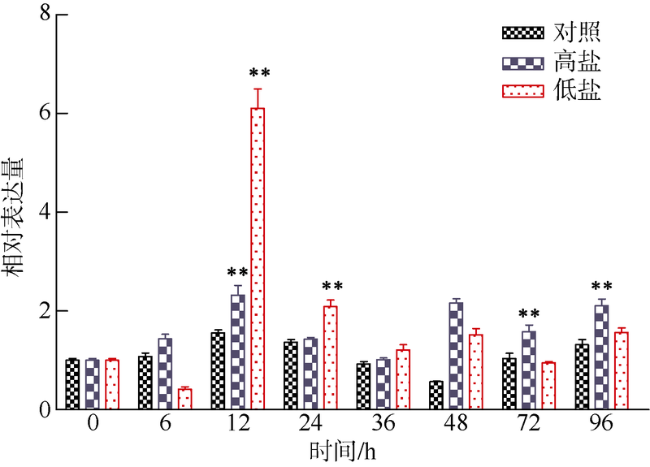
Fig. 6 Expression profiles of ChCommd1 in gill tissues under high salinity stress and low salinity stress图6 高盐和低盐胁迫下ChCommd1在腮组织中的表达模式 |
Fig. 7 Expression profiles of ChCommd1 in hemocytes under high salinity stress and low salinity stress图7 高盐和低盐胁迫下ChCommd1在血淋巴中的表达模式 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}