
Journal of Tropical Oceanography >
Seasonal variations of chlorophyll a and primary production and their influencing factors in the Pearl River Estuary
Received date: 2016-04-01
Request revised date: 2016-05-26
Online published: 2017-01-19
Supported by
Special Fund for Agro-scientific Research in the Public Interest (201403008)
National Natural Science Foundation of China (41130855, 31670458)
Strategic Priority Research Program of the Chinese Academy of Sciences (XDA11020305)
Copyright
Based on in situ physical and biological data collected in different seasons between 2014 and 2015, we described systematically the spatial-seasonal characteristics of chlorophyll a (Chl a) and primary production (PP) in the Pearl River Estuary (PRE) and their relationships with environmental factors. The results indicated that the average values of Chl a concentration and PP in the PRE were 3.77 mg·m-3 and 27.86 mg C·m-3·h-1, respectively. Both were higher in spring than in the other seasons, and the lowest were in winter. The spatial distribution of phytoplankton was mainly driven by the Pearl River discharge and the phytoplankton bloom was related to the location of salinity front. Due to nutrients limitation in the lower parts of the estuary, the bloom occurred in the upper parts when river discharge was relatively small. In contrast, although the nutrients were favorable for phytoplankton growth during wet season with large river discharge, the bloom was absent in the waters near the outlets likely associated with light limitation and dilution. The distribution of primary production in the PRE was well related to Chl a. The results also suggested that phosphorus was the potential limiting factor for phytoplankton growth in the lower parts of the PRE during wet season and low temperature was another reason for low primary production besides light in winter. Compared with previous results, the primary production was in the intermediate level in PRE during 2014~2015. The mean value of carbon assimilation index (AI) in the PRE was 7.51mg C·(mg Chl a)-1·h-1 and the carbon fixation level was 261.52 g C·m-2·y-1.
LIU Huajian , HUANG Liangmin , TAN Yehui , KE Zhixin , LIU Jiaxing , ZHAO Chunyu , WANG Junxing . Seasonal variations of chlorophyll a and primary production and their influencing factors in the Pearl River Estuary[J]. Journal of Tropical Oceanography, 2017 , 36(1) : 81 -91 . DOI: 10.11978/2016033
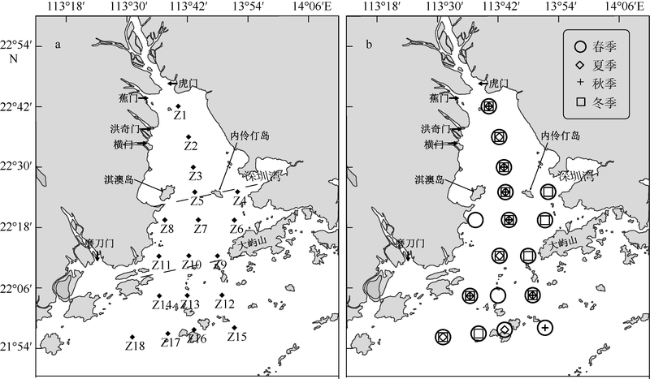
Fig. 1 Map of sampling stations (a: chlorophyll a; b: primary production) in the Pearl River Estuary (PRE)图1 珠江口站位分布图 |
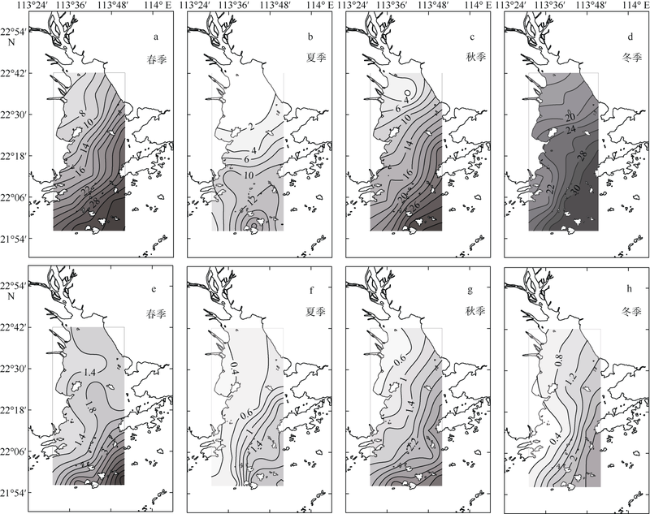
Fig. 2 Horizontal and seasonal variability of salinity (units: ‰; a~d) and transparency (units: m; e~h) in the PRE图2 珠江口表层盐度(单位: ‰; a~d)和透明度(单位: m; e~h)的时空变化 |
Tab. 1 The environmental parameters (mean ± SD) of surface water in the PRE during 2014—2015表1 2014—2015年珠江口表层环境参数 |
| 参数 | 春季(n = 18) | 夏季(n = 18) | 秋季(n = 18) | 冬季(n = 18) |
|---|---|---|---|---|
| 盐度/‰ | 21.26 ± 8.82 | 7.61 ± 5.70 | 18.40 ± 8.46 | 25.31 ± 5.68 |
| 温度/℃ | 20.99 ± 1.02 | 29.60 ± 0.80 | 25.75 ± 0.55 | 16.16 ± 0.82 |
| 透明度/ m | 2.27 ± 1.29 | 0.93 ± 0.57 | 2.13 ± 1.17 | 1.36 ± 0.89 |
| NO3- -N / (μmol·L-1) | 65.02 ± 36.00 | 76.69 ± 21.11 | 62.49 ± 36.47 | 34.36 ± 30.92 |
| NO2--N / (μmol·L-1) | 5.48 ± 3.75 | 13.79 ± 5.73 | 7.68 ± 4.31 | 4.23 ± 4.16 |
| NH4+-N/ (μmol·L-1) | 8.45 ± 7.09 | 1.02 ± 1.09 | 3.04 ± 1.36 | 18.10 ± 18.18 |
| DIN / (μmol·L-1) | 78.96 ± 44.40 | 91.51 ± 25.00 | 73.20 ± 37.70 | 56.68 ± 51.67 |
| PO43--P / (μmol·L-1) | 0.30 ± 0.19 | 0.72 ± 0.62 | 0.98 ± 0.28 | 0.62 ± 0.39 |
| SiO32--Si / (μmol·L-1) | 29.98 ± 19.31 | 86.72 ± 36.04 | 98.57 ± 38.20 | 30.54 ± 24.68 |
| N:P | 301 ± 183 | 162 ± 135a | 71 ± 32 | 84 ± 52 |
注: a表示n=12, 珠江口外部分站位PO43-浓度低于检测限。 |
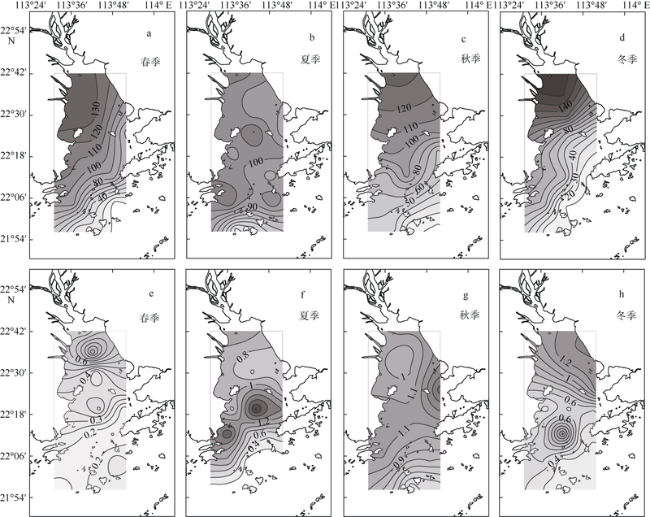
Fig. 3 Horizontal and seasonal variability of dissolved inorganic nitrogen (units: µmol·L-1; a~d) and phosphorus concentrations (units: µmol·L-1; e~h) in the PRE图3 珠江口表层溶解无机氮(单位: µmol·L-1; a~d)和磷酸盐(单位: µmol·L-1; e~h)浓度的时空变化 |
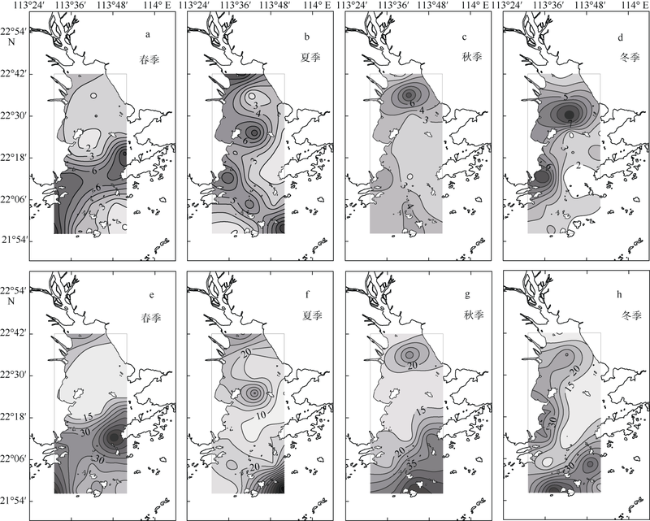
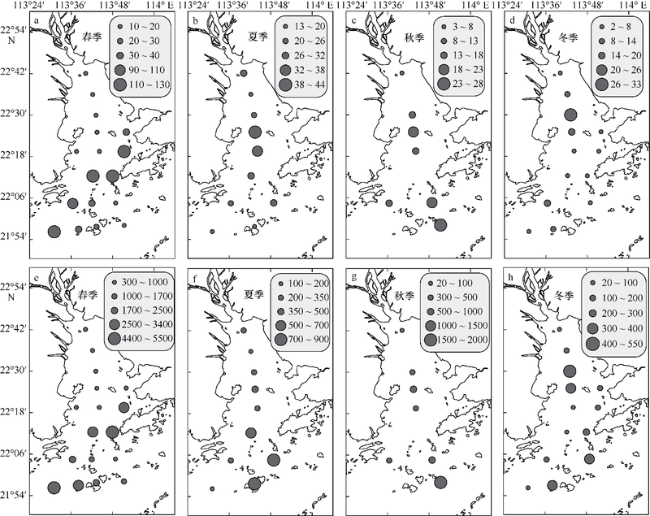
Fig. 4 Horizontal and seasonal variability of chlorophyll a (units: mg·m-3; a~d ) and integrated chlorophyll a (units: mg·m-2; e~h) in the PRE图4 珠江口表层叶绿素a (单位: mg·m-3; a~d)和水柱叶绿素a (单位: mg·m-2; e~h) 的时空变化 |
Fig. 5 Horizontal and seasonal variability of primary production (units: mg C·m-3·h-1; a~d) and integrated primary production (units: mg C·m-2·d-1; e~h) in the PRE图5 珠江口表层初级生产力(单位: mg C·m-3·h-1; a~d)与水柱初级生产力(单位: mg C·m-2·d-1; e~h) 的时空变化 |
Tab. 2 The carbon assimilation index [mg C·(mg Chl a) -1·h-1 ] of phytoplankton ( mean ± SD ) in the PRE表2 珠江口表层浮游植物碳同化系数[单位: mg C·(mg Chl a) -1·h-1 ] |
| 海区 | 春季 | 夏季 | 秋季 | 冬季 | 年平均 |
|---|---|---|---|---|---|
| 内伶仃洋 | 4.77 ± 0.65 | 5.55 ± 2.81 | 6.05 ± 2.75 | 4.59 ± 2.80 | 5.16 ± 2.15 |
| 外伶仃洋 | 11.79 ± 4.13 | 11.72 ± 2.37 | 6.31 | 4.17 ± 3.40 | 8.88 ± 5.20 |
| 珠江口外 | 10.70 ± 2.69 | 8.74 ± 3.05 | 7.61 ± 2.41 | 3.32 ± 1.26 | 8.37 ± 3.67 |
| 海区平均 | 9.65 ± 4.22 a | 8.06 ± 3.51 b | 6.75 ± 2.26 c | 4.04 ± 2.55 d | 7.51 ± 4.09 |
注: a表示n=17, b 表示n=10, c 表示n=7, d 表示n=13。 |
Tab. 3 Pearson correlation analysis for Chl a concentration, PP , IPP and environmental factors表3 叶绿素a (Chl a)浓度、初级生产力(PP)和水柱初级生产力(IPP)与环境因子之间的Pearson相关性 |
| 季节 | 参数 | 盐度 | 温度 | 透明度 | 硝酸盐 | DIN | 硅酸盐 | 磷酸盐 | 氮磷比 |
|---|---|---|---|---|---|---|---|---|---|
| 春季 | Chl a | -0.059 | -0.148 | -0.353 | 0.261 | 0.144 | 0.131 | -0.514* | 0.791** |
| PP | 0.225 | 0.081 | -0.259 | 0.058 | -0.054 | -0.078 | -0.582* | 0.750** | |
| 夏季 | IPP | 0.793** | -0.318 | 0.851** | -0.308 | -0.231 | -0.750* | -0.338 | 0.654 |
| 秋季 | PP | 0.768* | 0.723 | 0.684 | -0.719 | -0.696 | -0.784* | -0.602 | -0.638 |
| IPP | 0.926** | 0.678 | 0.995** | -0.866* | -0.873* | -0.905** | -0.960** | -0.836* | |
| 冬季 | Chl a | -0.484* | -0.155 | -0.092 | 0.488* | 0.402 | 0.382 | 0.145 | 0.464 |
注: *相关的显著性水平: p<0.05, **相关的显著性水平: p<0.01。 |
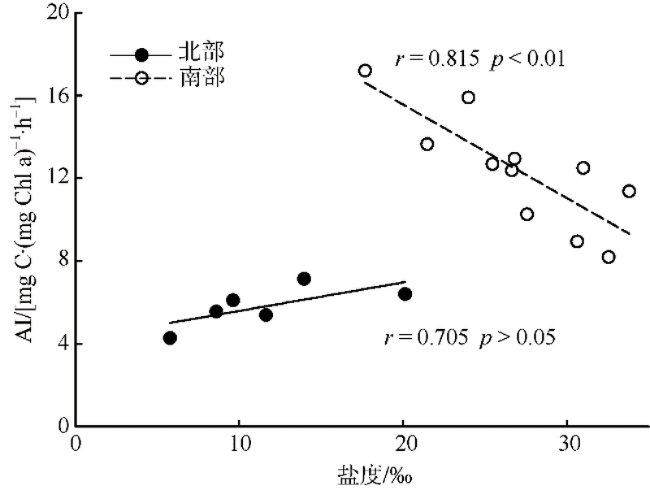
Fig. 6 Correlation between assimilation index and salinity图6 春季碳同化系数与盐度的关系 |
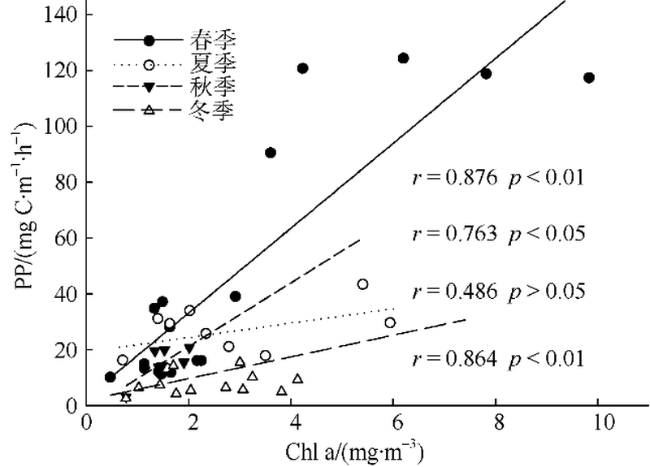
Fig. 7 Correlation between chlorophyll a and primary production图7 叶绿素a与初级生产力的关系 |
Tab. 4 The historical observations of phytoplankton primary production in the PRE表4 珠江口浮游植物初级生产力研究结果比较 |
| 时间 | 海区 | IPP/ (mg C·m-2·d-1 ) | 参考文献 |
|---|---|---|---|
| 1987—1993年 | 虎门至桂山岛 | 309 (夏86, 冬609) | 黄良民 等, 1997 |
| 1996—1997年 | 虎门上游至万山 | 685 (夏286, 冬1083) | 黄邦钦 等, 2005 |
| 1996—1997年 | 虎门上游至近海 | 134 (夏198, 冬69) | 蔡昱明 等, 2002 |
| 1999年7月 | 虎门至万山 | 100~400 | Yin et al, 2004b |
| 2006年 | 香港附近 | ~1300 | Ho et al, 2010 |
| 2012年6月 | 珠江口 | ~500 | Ye et al, 2015 |
| 2014—2015年 a | 虎门至万山 | 431 | 本研究 |
| 2014—2015年 b | 虎门至万山 | 716 | 本研究 |
注: ~表示估算值; a表示去除春季生产力极高值站位平均; b表示海区平均。 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}