
Journal of Tropical Oceanography >
Effect of aluminum on the growth and nitrogen fixation of a marine nitrogen-fixing cyanobacterium: Crocosphaera watsonii
Received date: 2016-07-25
Request revised date: 2016-08-31
Online published: 2017-04-06
Supported by
National Program on Key Basic Research Project (“973” Program)(2015CB4529000, 2015CB452903)
National Science Foundation of China (41276162, 41506150)
Strategic Priority Research Program of the Chinese Academy of Sciences (XDA11020200)
Special Fund for Agro-scientific Research in the Public Interest (201403008)
Science and Technology Planning Project of Guangdong Province (2014B030301064)
Copyright
Although a large amount of aluminum enters the ocean through atmospheric dust deposition every year, the effect of aluminum on marine phytoplankton is seldom examined. Few studies focused on the effects of aluminum on marine nitrogen-fixing cyanobacteria, which play important roles in nitrogen and carbon cycling in the ocean. In this study, a unicellular marine nitrogen-fixing cyanobacterium Crocosphaera watsonii grew in Aquil medium without silicate and nitrogen by semi-continuous cultures. We studied the effect of aluminum (0, 0.2, 2 and 20 μmol•L-1) on the growth rate and nitrogen fixation rate of Crocosphaera watsonii by using flow cytometry and 15N isotope tracer method. Results showed that aluminum inhibited the growth and nitrogen fixation of Crocosphaera watsonii. Results also showed that cell size, cellular pigments and photochemical efficiency of photosystem II (Fv/Fm) of Crocosphaera watsonii treated with aluminum were significantly decreased. These results demonstrate that aluminum has an inhibitory effect on the growth and nitrogen fixation of Crocosphaera watsonii under the condition of sufficient phosphate and trace nutrients.
LIU Jiaxing , ZHOU Linbin , KE Zhixin , LI Gang , SHI Rongjun , TAN Yehui . Effect of aluminum on the growth and nitrogen fixation of a marine nitrogen-fixing cyanobacterium: Crocosphaera watsonii[J]. Journal of Tropical Oceanography, 2017 , 36(2) : 12 -18 . DOI: 10.11978/2016069
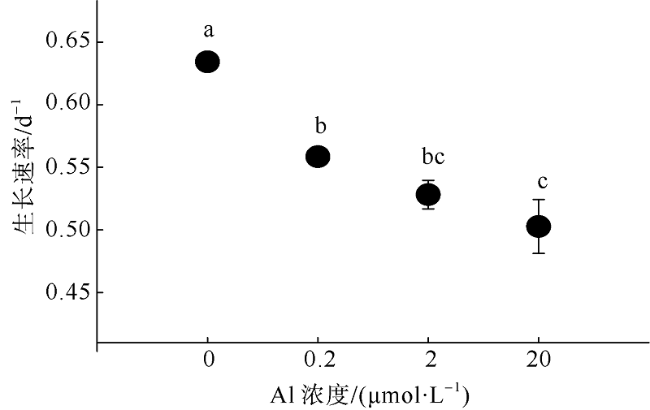
Fig. 1 Effect of aluminium on the growth rate of Crocosphaera watsonii. Different letters represent statistically significant differences at p < 0.05图1 铝对Crocosphaera watsonii生长速率的影响 |
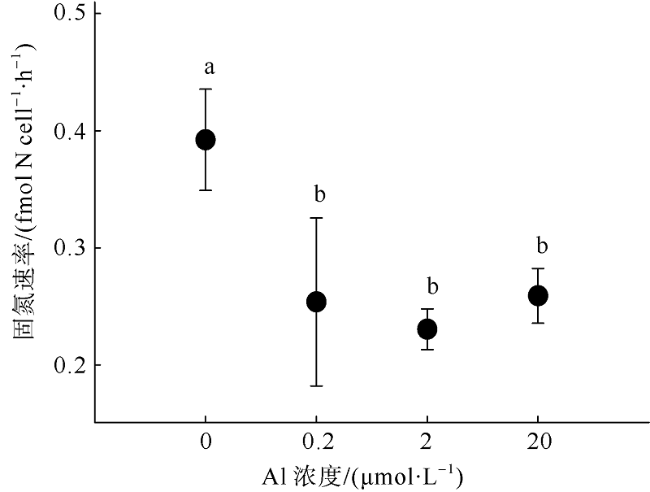
Fig. 2 Effect of aluminium on the nitrogen fixation rate of Crocosphaera watsonii图2 铝对Crocosphaera watsonii固氮速率的影响 |
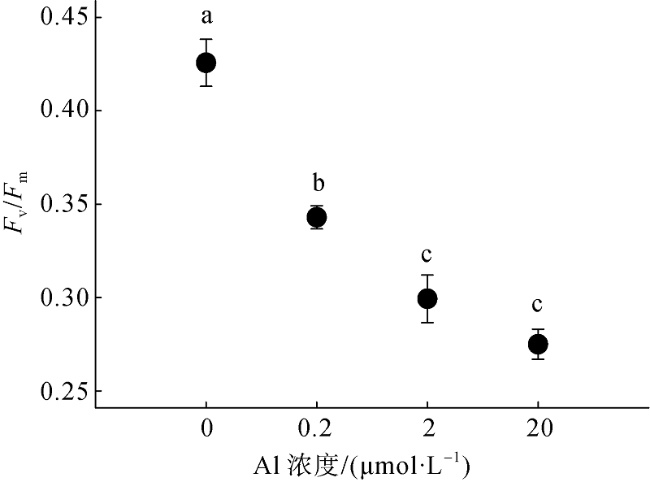
Fig. 3 Effect of aluminium on Fv/Fm of Crocosphaera watsonii图3 铝对Crocosphaera watsonii Fv/Fm比值的影响 |
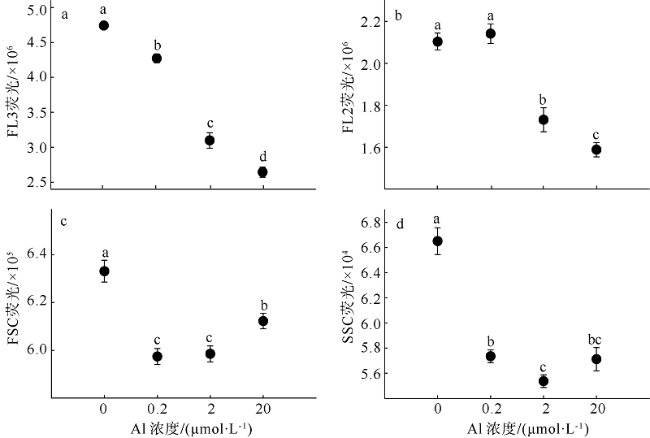
Fig. 4 Effect of aluminium on FL3 (a), FL2 (b), FSC (c), and SSC fluorescence (d) of Crocosphaera watsonii图4 铝处理后Crocosphaera watsonii细胞FL3(a)、FL2(b)、FSC(c)和SSC(d)荧光的变化 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}