
Journal of Tropical Oceanography >
Distribution characteristics of phytoplankton size structure in the western South China Sea in summer
Received date: 2016-10-21
Request revised date: 2016-11-23
Online published: 2017-07-26
Supported by
National Natural Sciences Foundation of China (41430968)
Collaborative Innovation Center for 21st-Century Maritime Silk Road Studies (2015HS05)
Dragon Program 4 (10689)
Copyright
Driven by the southwest monsoon, an offshore jet is usually formed in the western South China Sea (SCS) and sandwiched by a cyclonic eddy in the north and an anticyclonic eddy in the south, which affects the ecosystem of the region. Using in-situ and satellite data in September 2014, we analyzed the impact of this jet with two eddies on phytoplankton size structure in this region. The data showed that picophytoplankton (0.2~2 µm) dominated the surface, averaged at 76.7% of the total chlorophyll. The contributions of nanophytoplankton (2~20 µm) and microphytoplankton(20~200 µm)in the jet area were respectively higher, and had a positive relationship with the total chlorophyll. Comparatively higher percentage of microphytoplankton appeared in the anticyclonic eddy at surface (average 10.3%) than in the cyclonic eddy (average 3.6%). The results suggested that physical processes significantly influenced summertime surface phytoplankton size structure in the western South China Sea. Both the jet and eddies can affect phytoplankton size structure by increasing the contribution of microphytoplankton. Surface horizontal advection of phytoplankton by the northeastward jet formed the coastal upwelling area and was the main source of microphytoplankton in the open sea. The interactions of convergence and divergence in the eddies with the jet formed a chlorophyll front and increased the microphytoplankton component. Upwelling in the center of the cyclonic eddy brought up nutrients, which increased the percentage of the microphytoplankton component.
Key words: western South China Sea; phytoplankton size structure; coastal upwelling; jet; eddy
LIANG Wenzhao , TANG Danling . Distribution characteristics of phytoplankton size structure in the western South China Sea in summer[J]. Journal of Tropical Oceanography, 2017 , 36(4) : 93 -101 . DOI: 10.11978/2016104
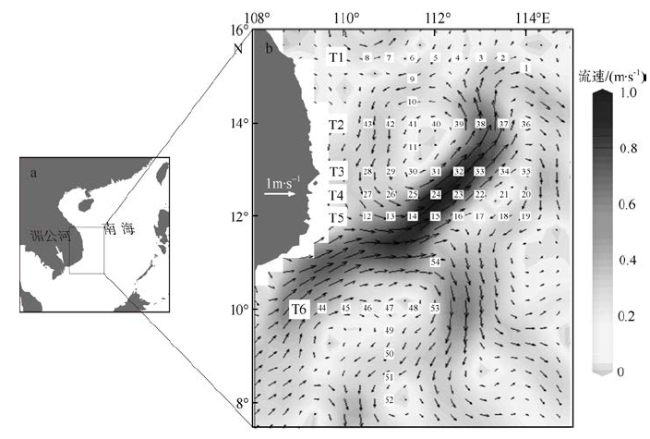
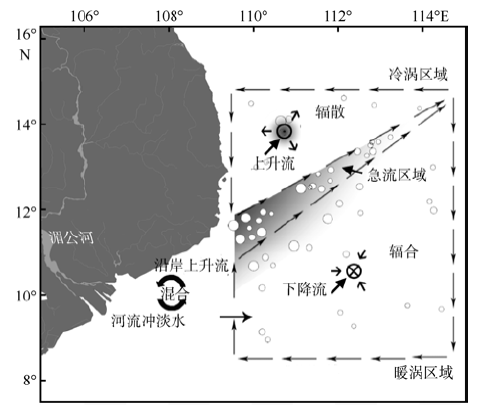
Fig. 1 The geographic location of the study region in the western South China Sea (a) and sampling stations, transects together with sea surface current in the study period (b)图1 研究区域的地理位置(a)及观测站点、断面及海表流场(b) |
Fig. 2 Sea level anomaly, temperature, salinity, and chlorophyll a concentration of the surface water in the western South China Sea from July to October in 2014图2 2014年7月至10月南海西部海洋表面高度异常、温度、盐度和叶绿素a浓度分布 |
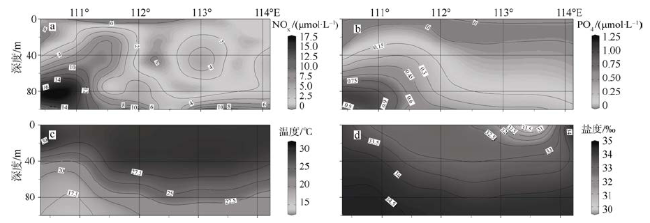
Fig. 3 The vertical sections of nitrite+nitrate (a), phosphate (b), temperature (c) and salinity (d) in the upper 100 m along transect T5 (12°N)图3 断面T5(12°N)的垂直剖面硝酸盐+亚硝酸盐(a)、磷酸盐(b)、温度(c)和盐度(d)分布图 |
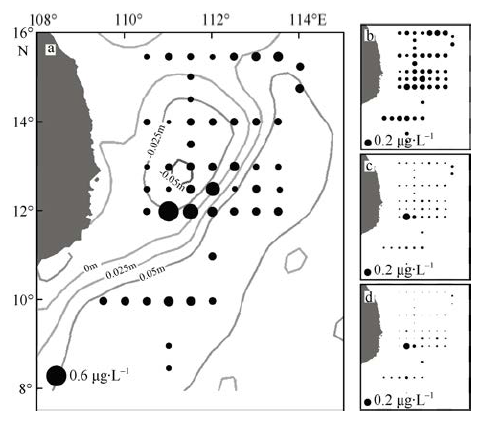
Fig. 4 Distribution of total chlorophyll a concentration together with SLA (Sep. 2014) (a). Surface concentrations of picophytoplankton (b), nanophytoplankton (c) and microphytoplankton (d) during the in situ observation图4 现场观测期间的总叶绿素a浓度及2014年9月海表高度异常卫星数据(a)以及微微型浮游植物(b)、微型浮游植物(c)和小型浮游植物(d)浓度分布 |
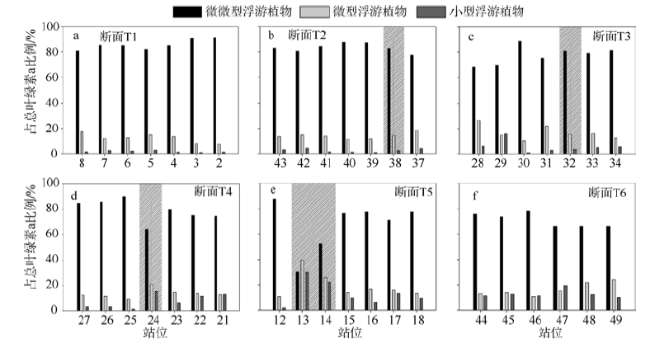
Fig. 5 Percentage contribution of various size fractions of phytoplankton to the total chlorophyll in the surface of T1(a), T2(b), T3(c), T4 (d), T5(e), and T6 (f)图5 断面T1(a)、T2(b)、T3(c)、T4(d)、T5(e)、T6(f)上各站点表层不同粒径大小占总叶绿素a的比例灰色背景表示急流区域 |
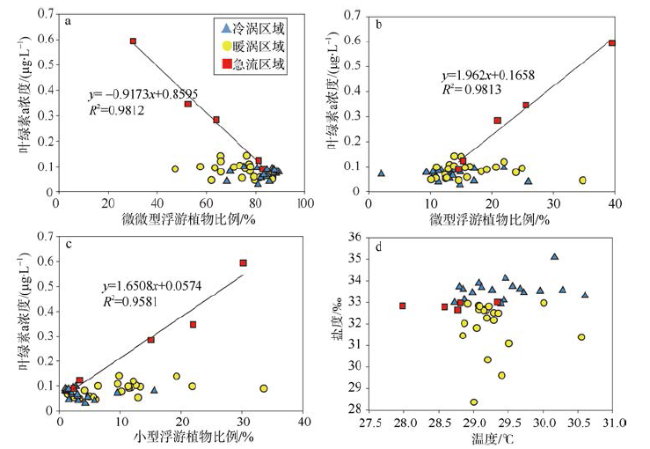
Fig. 6 Correlations between in situ sea surface total chlorophyll a and percentage of (a) picophytoplankton, (b) nanophytoplankton or (c) microphytoplankton. (d) Correlation between in situ surface temperature and salinity图6 叶绿素a浓度与微微型浮游植物的比例(a)、微型浮游植物的比例(b)和小型浮游植物的比例(c)的相关性, 以及温度与盐度的相关性(d) |
Fig. 7 Conceptual sea surface size structure characteristics of phytoplankton and its influence mechanism in summertime in the western South China Sea图7 南海夏季海表浮游植物粒径结构及影响机制概念图 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}