
Journal of Tropical Oceanography >
Studies on intraspecies diversity of Pseudo-nitzschia pungens from Chinese coastal waters*
Received date: 2017-02-15
Request revised date: 2017-05-26
Online published: 2018-02-02
Supported by
National Natural Science Foundation of China (31570205, 31370235)
Science and Technology Planning Project of Guangzhou, China (201607010370)
Copyright
To declare intraspecies diversity of Pseudo-nitzschia pungens Hasle, a total of 32 monoclonal strains of P. pungens from Chinese coastal waters were isolated and established. These strains were identified carefully, based on the morphology under both light microscopy and transmission electron microscopy, and on the molecular analysis inferred from the internal transcribed spacer region sequences. Two varieties were identified: P. pungens var. pungens (Grunow) Hasle and P. pungens var. averiensis Lundholm, Churro, Carreira, and Calado. Their morphologies were described and compared with allied taxa. Meanwhile, domoic acid (DA) was detected using a liquid chromatography tandem mass spectrometry (LC-MS/MS) injection, but no detectable DA was found. The results may provide detailed information for further studies of toxin production of Chinese Pseudo-nitzschia strains.
DONG Huanchang , HUANG Chunxiu , XU Guoshuang , LI Yang . Studies on intraspecies diversity of Pseudo-nitzschia pungens from Chinese coastal waters*[J]. Journal of Tropical Oceanography, 2018 , 37(1) : 12 -19 . DOI: 10.11978/2017017
Tab. 1 List of P. pungens monoclonal strains established in this study表1 本文建立的尖刺拟菱形藻单克隆培养株系 |
| 株系 | 分离地点 | 采样时间 | 种名 |
|---|---|---|---|
| MC215 | 珠海 | 2013年5月 | P. pungens var. pungens |
| MC218 | 葫芦岛 | 2013年8月 | P. pungens var. pungens |
| MC241 | 惠州 | 2013年10月 | P. pungens var. averiensis |
| MC255 | 厦门 | 2013年11月 | P. pungens var. averiensis |
| MC263 | 青岛 | 2013年11月 | P. pungens var. averiensis |
| MC289 | 青岛 | 2014年2月 | P. pungens var. pungens |
| MC290 | 青岛 | 2014年2月 | P. pungens var. pungens |
| MC295 | 惠州 | 2014年4月 | P. pungens var. pungens |
| MC404 | 惠州 | 2014年7月 | P. pungens var. averiensis |
| MC830 | 温州 | 2015年5月 | P. pungens var. pungens |
| MC831 | 温州 | 2015年5月 | P. pungens var. pungens |
| MC841 | 香港 | 2015年7月 | P. pungens var. averiensis |
| MC864 | 宁波 | 2015年8月 | P. pungens var. averiensis |
| MC867 | 宁波 | 2015年8月 | P. pungens var. averiensis |
| MC881 | 湛江 | 2015年8月 | P. pungens var. averiensis |
| MC888 | 湛江 | 2015年8月 | P. pungens var. averiensis |
| MC899 | 青岛 | 2015年9月 | P. pungens var. averiensis |
| MC915 | 深圳 | 2015年9月 | P. pungens var. averiensis |
| MC988 | 台湾海峡 | 2016年5月 | P. pungens var. pungens |
| MC3052 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3053 | 珠海 | 2016年5月 | P. pungens var. averiensis |
| MC3054 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3057 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3058 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3059 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3060 | 珠海 | 2016年5月 | P. pungens var. averiensis |
| MC3061 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3062 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3065 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3067 | 珠海 | 2016年5月 | P. pungens var. pungens |
| MC3119 | 黄海 N:122°50′, E:32°00′ | 2016年7月 | P. pungens var. pungens |
| MC3195 | 台湾海峡 | 2016年8月 | P. pungens var. averiensis |
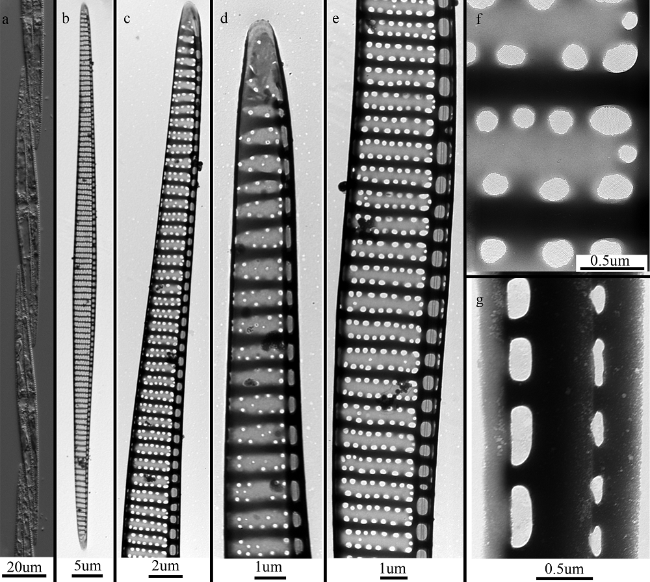
Fig. 1 Pseudo-nitzschia pungens var. pungens. (a) LM, chain colony; (b) shape of whole valve; (c) valve end; (d) valve end; (e) middle of valve; (f) detail of poroids; (g) bands. b’g: TEM图1 尖刺拟菱形藻原变种 |
Fig. 2 Pseudo-nitzschia pungens var. averiensis. (a) LM, chain colony; (b) shape of whole valve; (c) valve end; (d) middle of valve; e~h: bands; b~h: TEM图2 尖刺拟菱形藻阿维罗变种 |
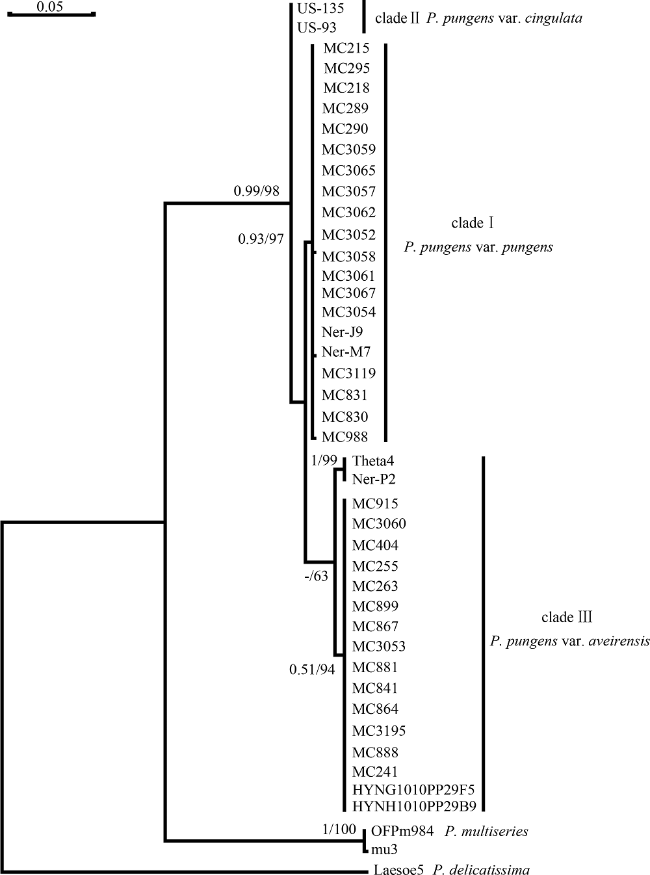
Fig. 3 Molecular phylogenetic tree based on ITS1-5.8S-ITS2 rDNA with Pseudo-nitzschia delicatissima as outgroups图3 基于核糖体转录间隔区ITS1-5.8S-ITS2的分子系统树 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}