
Journal of Tropical Oceanography >
Subsurface chlorophyll a maximum and its possible causes in the southern South China Sea*
Received date: 2017-02-23
Request revised date: 2017-04-24
Online published: 2018-02-02
Supported by
National Key Research and Development Program of China (2016YFC1401604)
Open Program of Key Laboratory of South China Sea Fishery Resources Development and Utilization, Ministry of Agriculture (LSF2012-03)
Financial Fund of Ministry of Agriculture (NFZX2013)
Copyright
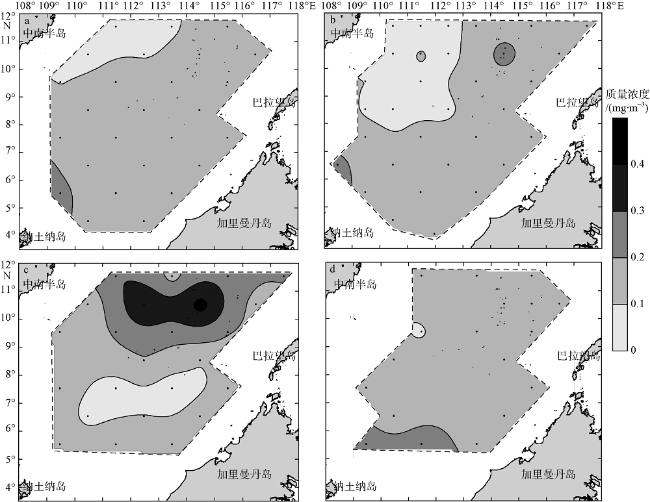
The four-season environment ecological comprehensive surveys were carried out at 34 stations in the southern South China Sea by the research vessel “NanFeng” from March to December in 2013. Results show that the average chlorophyll a (Chl a) concentration of shallow water at 150 m in early spring (0.14 mg•m-3) and early summer (0.12 mg•m-3) were lower than that in early autumn (0.18 mg•m-3) and early winter (0.16 mg•m-3). This was mainly due to the smallest spring wind speed in early spring and the highest water temperature in early summer, which were not conducive to vertical mixing, limiting deep nutrients being brought up. The subsurface Chl a maximum in the four seasons all appeared at both 50 m (0.24~0.26 mg•m-3) or 75 m (0.22~0.26 mg•m-3), and the difference of Chl a concentration at these two depths was small, with little season change. The 50 m and 75 m layers were located in the middle part and near the bottom of the euphotic seawater, and the light received by the two layers can meet the growth need of phytoplankton. With the combined impact of mixed layer depth and the upper bound of the thermocline, the Chl a concentration in the 50 m layer was mainly controlled by the supply of nutrients, and that in the 75 m layer, by ocean temperature, both being important factors affecting phytoplankton growth.
LIAO Xiuli , DAI Ming , GONG Xiuyu , LIU Huaxue , HUANG Honghui . Subsurface chlorophyll a maximum and its possible causes in the southern South China Sea*[J]. Journal of Tropical Oceanography, 2018 , 37(1) : 45 -56 . DOI: 10.11978/2017020
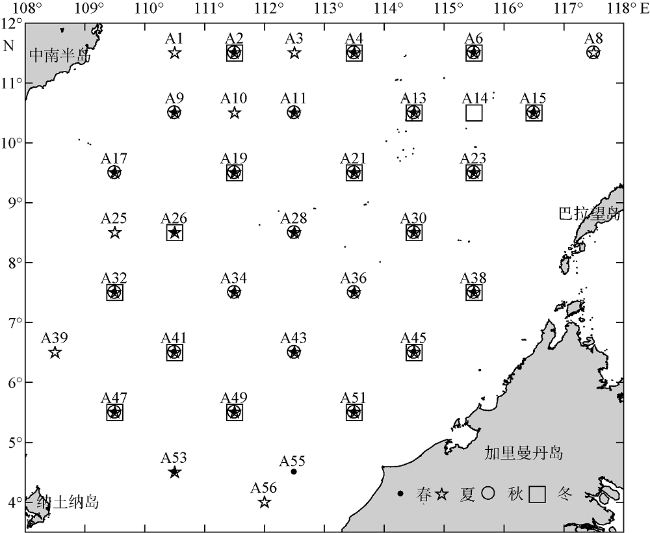
Fig. 1 Sampling sites in the southern South China Sea. The dots, stars, circles, and squares represent sampling sites in spring, summer, autumn, and winter, respectively图1 南海南部采样站位图 |
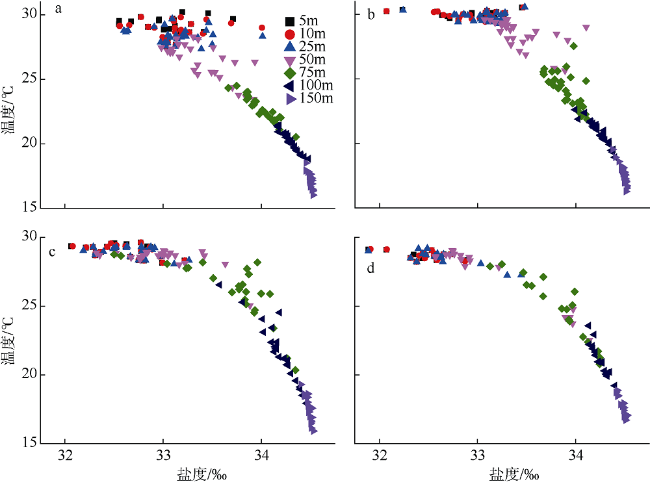
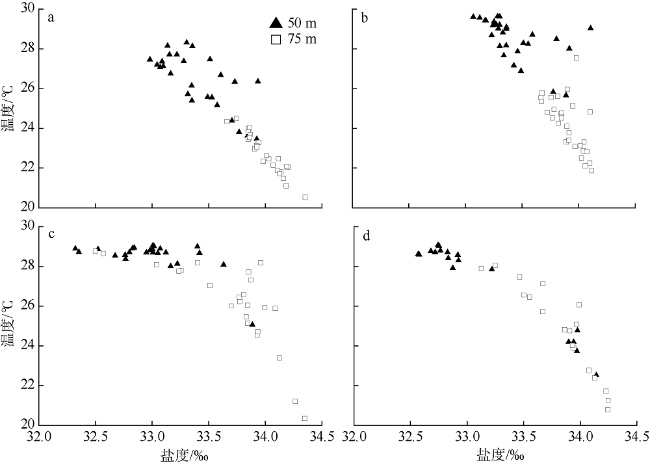
Fig. 2 Scatter diagrams of vertical distributions of water temperature and salinity of the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively图2 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d)温盐垂直分布散点图 |
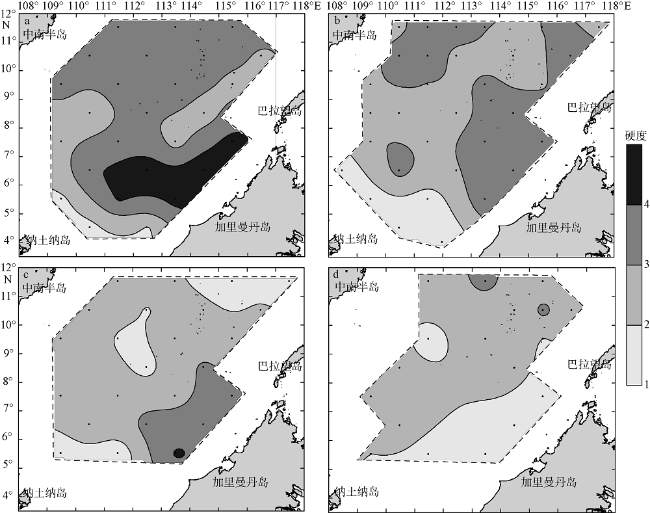
Fig. 3 Spatial distributions of the hardness of water column in the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively图3 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d)水柱硬度的空间分布 |
Fig. 4 Scatter diagrams of vertical distributions of Chl a in the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively. Units: mg•m-3图4 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d) Chl a质量浓度垂直分布散点图 |
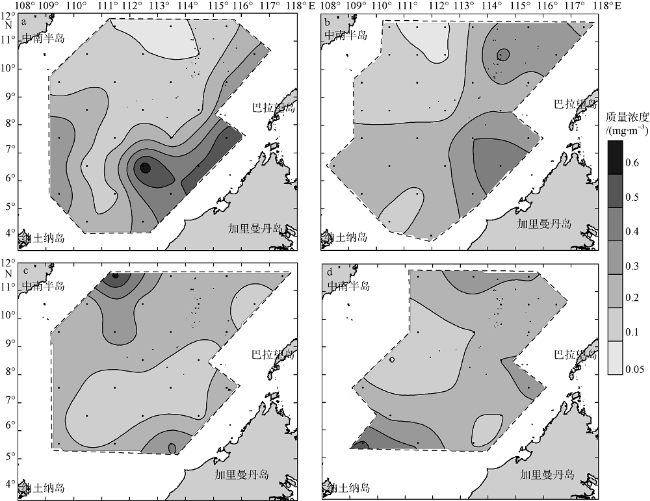
Fig. 5 Spatial distribution of average Chl a of water column in the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively. Units: mg•m-3图5 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d)水柱Chl a质量浓度均值的空间分布 |
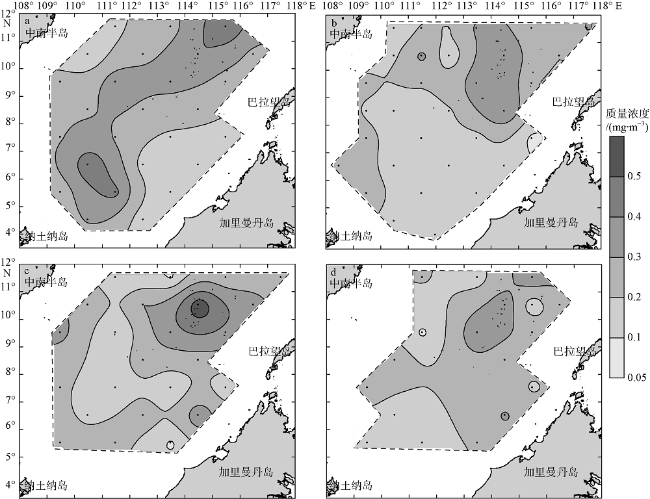
Fig. 6 Spatial distributions of Chl a at 50 m in the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively. Units: mg•m-3图6 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d) 50m层Chl a质量浓度的空间分布 |
Fig. 7 Spatial distributions of Chl a at 75 m in the four seasons. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively. Units: mg•m-3图7 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d) 75m层Chl a质量浓度的空间分布 |
Tab. 1 Correlation analysis between nutrients and Chl a in the subsurface chlorophyll maximum layer表1 次表层叶绿素最大值层的Chl a质量浓度与营养盐质量浓度的相关分析 |
| 水层 | 水温 | 盐度 | NO2-N | NO3-N | NH4-N | DIN | DIP | DIN/DIP | SiO3-N | 样本数 |
|---|---|---|---|---|---|---|---|---|---|---|
| 50m | -0.463** | 0.414** | 0.502** | 0.214* | -0.145 | -0.01 | 0.372** | -0.139 | 0.195 | 99 |
| 75m | 0.248* | -0.245* | 0.121 | -0.27* | -0.125 | -0.306** | -0.264* | -0.014 | -0.148 | 99 |
注: *: p<0.05; **: p<0.01 |
Fig. 8 Scatter plots of temperature and salinity at 50 and 75 m. (a), (b), (c), and (d) are early spring, early summer, early autumn, and early winter, respectively图8 南海南部海域早春(a)、初夏(b)、初秋(c)和初冬(d) 50m和75m层的温度、盐度散点图 |
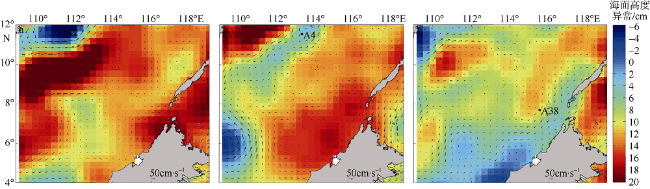
Fig. 9 Sea level anomalies and geostrophic circulation distribution maps. (a) 20130926; (b) 20131118; (c) 20130627图9 海面高度异常和地转流矢量分布图 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}