
Journal of Tropical Oceanography >
In situ feeding of copepodites in the water near Daya Bay nuclear power plant
Received date: 2017-05-21
Request revised date: 2017-08-14
Online published: 2018-04-11
Supported by
National Key Research and Development Project (2016YFC0502805)
Science and Technology Planning Projects of Guangdong Province, China (2015A020216013)
Strategic Priority Research Program of Chinese Academy of Sciences (XDA13020100)
National Basic Research Program of China (“973” Program) (2015CB452904)
National Natural Science Foundation of China (41276160)
Science and Technology Planning Project of Guangdong Province, China (2017B030314052)
Copyright
Copepods play important roles in maintaining the structure of marine ecosystem due to their key position in food chain and their numerous species, quantities and wide distribution. The nutrients obtained by copepod larvae can directly affect their development, then the complement of adult copepods and even their population stability. In this study, in situ feeding of copepodites was evaluated by molecular methods in the waters near the nuclear power plant in Daya Bay (S1: control station, S2: outfall station) in summer 2015. The results showed that 16 prey species in total were identified in both stations, including diatoms (11 species), tunicate (two species), fungi (one species), ichthyosporea (one species), and oomycetes (one species). Diatoms (47.30%) and tunicate (41.89%) were the most abundant food types revealed by the significant proportion of the clones. Similar feeding diversity was discovered in copepodites at S1 and S2, with nine prey species at S1 and 10 at S2, but significant differences in diet composition were revealed, with metazoan (61.54%) and diatoms (68.57%) dominating the diets of copepodites at S1 and S2, respectively. In addition, copepodites appeared to be more herbivorous at S2 with a lower Omnivory Index (0.31) than at S1 (0.72). These results suggested that copepodites could optimize their diet composition by consuming a reasonable percentage of animal and plant materials according to available food sources. More phytoplankton, especially diatoms as diet of copepodites at the outfall station implied that copepodites might change their feeding habits with global warming development.
Key words: Daya Bay; copepodites; feeding; diversity; increased temperature
XU Cuilian , LI Tao , HU Simin , WANG Youjun , HUANG Hui , LIU Sheng . In situ feeding of copepodites in the water near Daya Bay nuclear power plant[J]. Journal of Tropical Oceanography, 2018 , 37(2) : 17 -25 . DOI: 10.11978/2017058
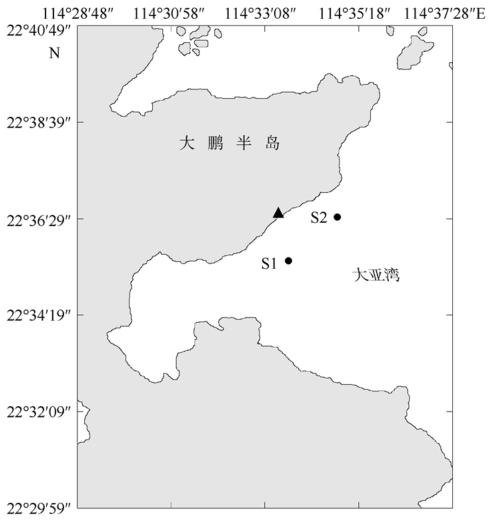
Fig. 1 Sampling sites in Daya Bay. Triangle is the location of Daya Bay Nuclear Power Plant and Ling-Ao Nuclear Power Plant. S1: control station; S2: outfall station图1 大亚湾调查和采样站位 |
Tab. 1 Diversity indices and biomass of phytoplankton表1 浮游植物多样性指数及生物量 |
| 种类数 | 丰度百分比 | 细胞丰度/ (×105cells·L-1) | 香农-威纳指数a | 优势种 | 叶绿素a含量/(μg·L-1) | |||
|---|---|---|---|---|---|---|---|---|
| 甲藻 | 硅藻 | 甲藻 | 硅藻 | |||||
| S1 | 7 | 14 | 66.14% | 38.36% | 4.38 | 1.89 | Scrippsiella trochoidea | 5.09 |
| S2 | 5 | 12 | 71.94% | 28.06% | 3.92 | 1.39 | 3.18 | |
注: a香农-威纳(Shannon-Weaver)指数\(H |
Tab. 2 Diet diversity indices of copepodites表2 桡足幼体现场食物多样性指数 |
| 样品编号 | 种类数 | 有效克隆数 | 辛普森指数b | 香农-威纳指数c | Chao1 指数d |
|---|---|---|---|---|---|
| S1 | 9 | 39 | 0.6772 | 1.593 | 10.5 |
| S2 | 10 | 35 | 0.8245 | 1.987 | 11 |
注: b辛普森(Simpson)指数\(D=1-\sum^s_{i=1}P^2_i\); Pi=Ni/N, 式中 S是得到的种类数量, Ni 是含有 i 条序列的序列数目, N是总序列的数量(Simpson, 1949)。c香农-威纳(Shannon-Weaver)指数的计算公式同 |
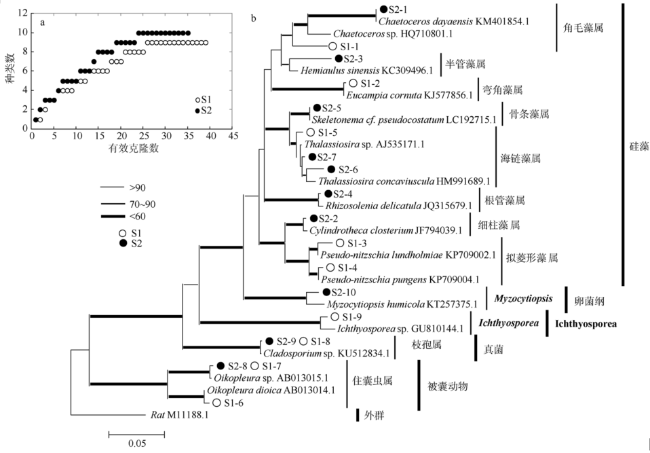
Fig. 2 Relationship between clones and taxon of copepodite diet (a) and neighbor joining tree of 18S rDNA sequences obtained from copepodite diet (b)图2 送测有效克隆样品数所覆盖的种类数(a)和桡足幼体克隆文库测的18S rDNA序列构建的邻接树(b) |
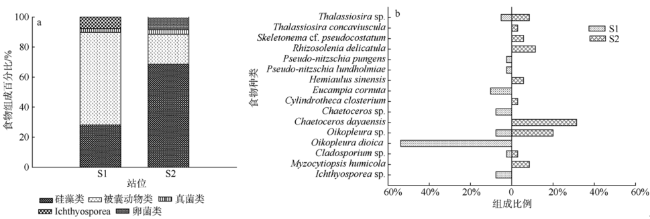
Fig. 3 Prey composition of copepodites. (a) group of prey composition; (b) species of prey composition图3 桡足幼体肠道食物组成 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.6-2007 海洋调查规范第6部分: 海洋生物调查[S]. 北京: 中国标准出版社.
General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2008. GB/T 12763.6-2007 Specifications for oceanographic survey—Part 6: Marine biological survey[S]. Beijing: China Standard Publishing House (in Chinese).
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
VAN DE WAAL D B,
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}