
Journal of Tropical Oceanography >
Spatiotemporal variations of virioplankton and bacterioplankton abundance in Sanya waters and their related environmental factors
Received date: 2017-05-26
Request revised date: 2017-09-27
Online published: 2018-04-11
Supported by
Knowledge Innovation Program of the Chinese Academy of Sciences (SIDSSE-201303)
The National Natural Science Foundation of China (41406180)
Copyright
Surface water samples were collected monthly for a total of 12 months from three stations in Sanya. Their virioplankton and bacterioplankton abundances were measured by flow cytometry and their correlationships with evnironmental factors were also studied. The results show that the abundance of virioplankton (7.63×106 viruses·mL-1 on average) was higher than bacterioplankton abundance (1.52×106 cells·mL-1 on average). Virioplankton and bacterioplankton abundances in the Estuary of Sanya River were significantly different from those on the west side of the Luhuitou Peninsula and Xiaodonghai. The highest virioplankton and bacterioplankton abundances appeared in the Estuary of Sanya River, with obvious seasonal variation(P<0.01). The abundances of virioplankton and bacterioplankton were significantly positively correlated (r = 0.800, P<0.01); and chlorophyll a, N nutrients (NO- 2, NO- 2+NO- 3, NH+ 4) are their key ecological factors (P < 0.01).
Key words: Sanya; virioplankton; bacterioplankton; flow cytometry
JING Hongmei , WEI Yu , ZHENG Liping , XU Qiutong , ZHANG Yue , ZHU Wenda . Spatiotemporal variations of virioplankton and bacterioplankton abundance in Sanya waters and their related environmental factors[J]. Journal of Tropical Oceanography, 2018 , 37(2) : 36 -46 . DOI: 10.11978/2017064
Fig. 1 Sampling stations off the Luhuitou Peninsula图1 鹿回头半岛近海采样站位 |
Fig. 2 Total abundance of bacterioplankton at the three stations图2 三个站位浮游细菌丰度 |
Tab. 1 Differences in total abundances of bacterioplankton and virioplankton at the three stations表1 三个站位浮游细菌丰度差异性 |
| 站位 | 细菌 | 病毒V-Ⅰ | 病毒V-Ⅱ | 总病毒 | ||||
|---|---|---|---|---|---|---|---|---|
| 鹿回头半岛西侧 | 小东海 | 鹿回头半岛西侧 | 小东海 | 鹿回头半岛西侧 | 小东海 | 鹿回头半岛西侧 | 小东海 | |
| 小东海 | 0.73 | 0.812 | 0.469 | 0.492 | ||||
| 三亚河口 | 0.009** | 0.008** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** |
注: *P<0.05; ** P<0.01。 |
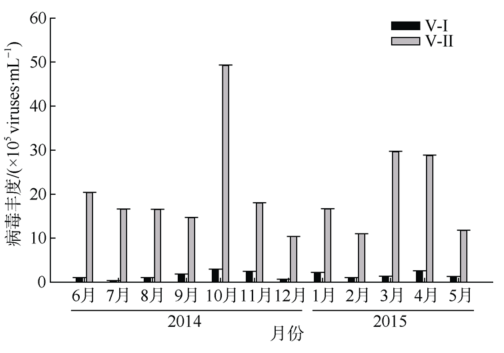
Fig. 3 Abundance of virioplankton on the west side of the Luhuitou Peninsula图3 鹿回头半岛西侧海域浮游病毒丰度 |
Fig. 4 Abundance of virioplankton in Xiaodonghai图4 小东海浮游病毒丰度 |
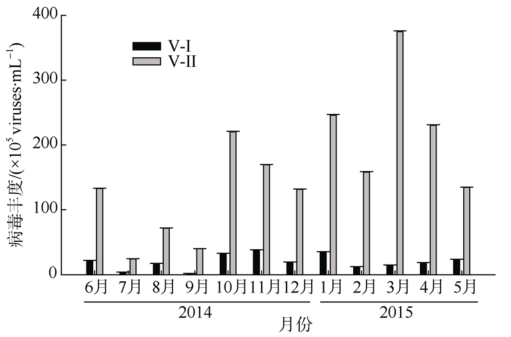
Fig. 5 Abundance of virioplankton in Sanya River Estuary图5 三亚河口浮游病毒丰度 |
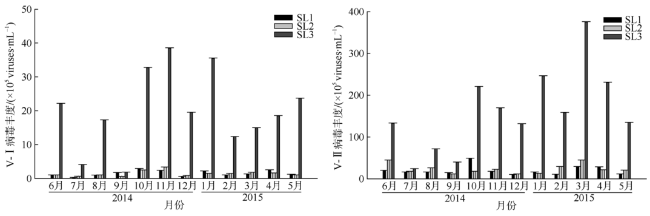
Fig. 6 Total abundance of low and high DNA virioplankton at the three stations图6 三个站位V-Ⅰ(左图)、V-Ⅱ(右图)病毒丰度 |
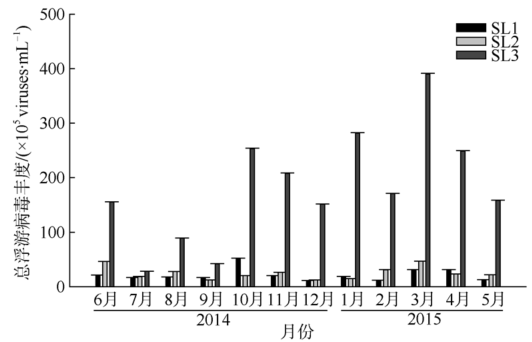
Fig. 7 Total abundance of virioplankton at the three stations图7 三个站位总浮游病毒丰度 |
Tab. 2 Pearson correlation coefficients between abundance of virioplankton, bacterioplankton and environmental parameters at station SL1表2 站位SL1浮游病毒丰度与浮游细菌、环境参数间的Pearson相关系数 |
| 相关影响因子 | Vir | Bac | VBR | Sal | Tem | pH | Chl a (>20μm) | Chl a (2~20μm) | Chl a (GF/F) | NO- 2 | NO- 2 +NO- 3 | PO3- 4 | SiO2- 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bac | -0.034 | ||||||||||||
| VBR | 0.632* | -0.549 | |||||||||||
| Sal | -0.310 | -0.337 | -0.068 | ||||||||||
| Tem | 0.039 | 0.668* | -0.396 | -0.826** | |||||||||
| pH | 0.395 | 0.287 | -0.267 | -0.221 | 0.371 | ||||||||
| Chl a (>20μm) | 0.637* | -0.192 | 0.163 | 0.267 | -0.335 | 0.507 | |||||||
| Chl a (2~20μm) | -0.052 | -0.424 | 0.060 | 0.724** | -0.769** | -0.134 | 0.571 | ||||||
| Chl a (GF/F) | 0.636* | -0.224 | 0.173 | 0.069 | -0.124 | 0.669* | 0.891** | 0.387 | |||||
| NO- 2 | -0.031 | 0.838** | -0.333 | -0.185 | 0.397 | 0.087 | -0.184 | -0.194 | -0.188 | ||||
| NO- 2+NO- 3 | 0.154 | 0.799** | -0.288 | -0.281 | 0.432 | 0.188 | 0.022 | -0.157 | 0.011 | 0.957** | |||
| PO3- 4 | 0.163 | -0.410 | 0.431 | 0.155 | -0.509 | -0.036 | 0.265 | 0.545 | 0.204 | -0.214 | -0.206 | ||
| SiO2- 3 | 0.049 | 0.686* | -0.230 | -0.070 | 0.191 | 0.040 | -0.001 | -0.013 | -0.057 | 0.942** | 0.949** | -0.062 | |
| NH+ 4 | 0.005 | 0.777** | -0.238 | -0.203 | 0.432 | 0.073 | -0.235 | -0.258 | -0.147 | 0.955** | 0.893** | -0.311 | 0.857** |
注: *P < 0.05; **P < 0.01。Vir: 浮游病毒丰度; Bac: 浮游细菌丰度; VBR: 病毒/细菌比值; Sal: 盐度; Tem: 温度; Chl a (>20μm): >20μm的Chl a浓度; Chl a (2~20μm): 2~20μm的Chl a浓度; Chl a (GF/F) : GF/F的Chl a浓度; NO- 2: 亚硝酸盐浓度; NO- 2+NO- 3: 硝酸盐和亚硝酸盐浓度; PO3- 4: 磷酸盐浓度; SiO2- 3: 硅酸盐浓度; NH+ 4: 铵盐浓度。下同。 |
Tab. 3 Pearson correlation coefficients between abundance of virioplankton, bacterioplankton and environmental parameters at station SL2表3 站位SL2浮游病毒丰度与浮游细菌、环境参数间的Pearson相关系数 |
| 相关影响因子 | Vir | Bac | VBR | Sal | Tem | pH | Chl a (>20μm) | Chl a (2~20μm) | Chl a (GF/F) | NO- 2 | NO- 2+NO- 3 | PO3- 4 | SiO2- 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bac | 0.332 | ||||||||||||
| VBR | 0.446 | -0.636* | |||||||||||
| Sal | -0.019 | -0.378 | 0.327 | ||||||||||
| Tem | 0.046 | 0.610* | -0.446 | -0.791** | |||||||||
| pH | -0.042 | 0.138 | -0.210 | 0.064 | 0.369 | ||||||||
| Chl a(>20μm) | 0.424 | -0.408 | 0.553 | 0.393 | -0.589* | -0.183 | |||||||
| Chl a(2~20μm) | -0.114 | -0.412 | 0.315 | 0.027 | 0.068 | 0.514 | 0.002 | ||||||
| Chl a(GF/F) | 0.278 | -0.422 | 0.438 | -0.015 | -0.146 | 0.318 | 0.605* | 0.601* | |||||
| NO- 2 | -0.491 | 0.046 | -0.499 | -0.486 | 0.348 | 0.255 | -0.433 | -0.103 | -0.112 | ||||
| NO- 2+NO- 3 | -0.247 | 0.241 | -0.525 | -0.541 | 0.450 | 0.023 | -0.488 | -0.241 | -0.311 | 0.613* | |||
| PO3- 4 | -0.285 | 0.148 | -0.311 | 0.406 | -0.193 | -0.083 | 0.157 | -0.364 | -0.347 | -0.18 | -0.318 | ||
| SiO2- 3 | -0.290 | 0.036 | -0.328 | 0.065 | -0.240 | -0.156 | -0.305 | -0.121 | -0.254 | 0.353 | 0.334 | -0.164 | |
| NH+ 4 | 0.638* | 0.492 | 0.123 | -0.074 | 0.253 | 0.281 | -0.116 | -0.052 | 0.057 | -0.079 | -0.303 | -0.188 | -0.200 |
Tab. 4 Pearson correlation coefficients between abundance of virioplankton, bacterioplankton and environmental parameters at station SL3表4 站位SL3浮游病毒丰度与浮游细菌、环境参数间的Pearson相关系数 |
| 相关影响因子 | Vir | Bac | VBR | Sal | Tem | pH | Chl a (>20μm) | Chl a (2~20μm) | Chl a (GF/F) | NO- 2 | NO- 2+NO- 3 | PO3- 4 | SiO2- 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bac | 0.677* | ||||||||||||
| VBR | 0.164 | -0.539 | |||||||||||
| Sal | 0.474 | 0.593* | -0.127 | ||||||||||
| Tem | -0.563 | -0.686* | 0.205 | -0.680* | |||||||||
| pH | 0.059 | -0.409 | 0.720** | -0.140 | 0.433 | ||||||||
| Chl a(>20μm) | 0.469 | 0.724** | -0.257 | 0.638* | -0.597* | -0.138 | |||||||
| Chl a(2~20μm) | 0.406 | 0.539 | -0.230 | 0.367 | -0.549 | -0.219 | 0.089 | ||||||
| Chl a(GF/F) | 0.456 | 0.564 | -0.167 | 0.394 | -0.316 | 0.047 | 0.312 | 0.759** | |||||
| NO- 2 | 0.531 | 0.452 | 0.159 | 0.351 | -0.719** | -0.143 | 0.220 | 0.633* | 0.250 | ||||
| NO- 2+NO- 3 | 0.109 | 0.078 | 0.153 | -0.132 | -0.517 | -0.196 | 0.001 | 0.266 | -0.171 | 0.776** | |||
| PO3- 4 | 0.403 | -0.018 | 0.528 | -0.002 | -0.327 | 0.233 | 0.091 | 0.298 | 0.130 | 0.488 | 0.449 | ||
| SiO2- 3 | -0.033 | -0.104 | 0.183 | -0.257 | -0.359 | -0.137 | -0.193 | 0.088 | -0.351 | 0.600* | 0.930** | 0.397 | |
| NH+ 4 | 0.277 | 0.167 | 0.203 | -0.016 | -0.521 | -0.074 | -0.033 | 0.604* | 0.181 | 0.851** | 0.837** | 0.712** | 0.738** |
Tab. 5 Pearson correlation coefficients between abundance of virioplankton, bacterioplankton and environmental parameters fo the overall level at the surveyed stations表5 调查海域总体水平浮游病毒丰度与浮游细菌、环境参数间的Pearson相关系数 |
| 相关影响因子 | Vir | Bac | VBR | Sal | Tem | pH | Chl a (>20μm) | Chl a (2~20μm) | Chl a (GF/F) | NO- 2 | NO- 2+NO- 3 | PO3- 4 | SiO2- 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bac | 0.800** | ||||||||||||
| VBR | 0.331* | -0.230 | |||||||||||
| Tem | -0.211 | -0.218 | -0.054 | -0.621** | |||||||||
| pH | -0.117 | -0.273 | 0.236 | -0.014 | 0.389* | ||||||||
| Chl a(>20μm) | 0.546** | 0.614** | 0.033 | 0.280 | -0.461** | -0.042 | |||||||
| Chl a(2~20μm) | 0.582** | 0.639** | -0.029 | 0.071 | -0.303 | -0.178 | 0.251 | ||||||
| Chl a(GF/F) | 0.689** | 0.683** | 0.087 | -0.002 | -0.180 | -0.006 | 0.503** | 0.804** | |||||
| NO- 2 | 0.792** | 0.697** | 0.275 | -0.147 | -0.235 | -0.187 | 0.379* | 0.700** | 0.556** | ||||
| NO- 2+NO- 3 | 0.587** | 0.497** | 0.225 | -0.405* | -0.086 | -0.173 | 0.254 | 0.462** | 0.315 | 0.887** | |||
| PO3- 4 | 0.134 | -0.010 | 0.244 | 0.137 | -0.250 | 0.049 | 0.088 | 0.133 | 0.030 | 0.143 | 0.086 | ||
| SiO2- 3 | 0.359* | 0.271 | 0.198 | -0.353* | -0.142 | -0.161 | 0.072 | 0.275 | 0.067 | 0.720** | 0.908** | 0.078 | |
| NH+ 4 | 0.714** | 0.571** | 0.312 | -0.335* | -0.146 | -0.162 | 0.271 | 0.677** | 0.536** | 0.938** | 0.905** | 0.204 | 0.771** |
Tab. 6 Comparison with existing studies about Sanya Bay表6 与已有的三亚湾研究进行比较 |
| 研究 | 细菌丰度/(×108cells·L-1) | 时间 | 站位 |
|---|---|---|---|
| 周伟华 等(2006) | 11.32±7.26 | 2004秋 | 三亚湾 |
| 周伟华 等(2007) | 6.70±2.49 | 2005春 | 三亚湾 |
| 8.41±2.51 | 2005夏 | 三亚湾 | |
| 5.78±2.81 | 2005秋 | 三亚湾 | |
| 8.84±4.67 | 2005冬 | 三亚湾 | |
| 本研究 | 6.56±5.00 | 2014—2015 | 鹿回头西侧 |
| 5.85±4.93 | 2014—2015 | 小东海 | |
| 33.3±29.19 | 2014—2015 | 三亚河口 |
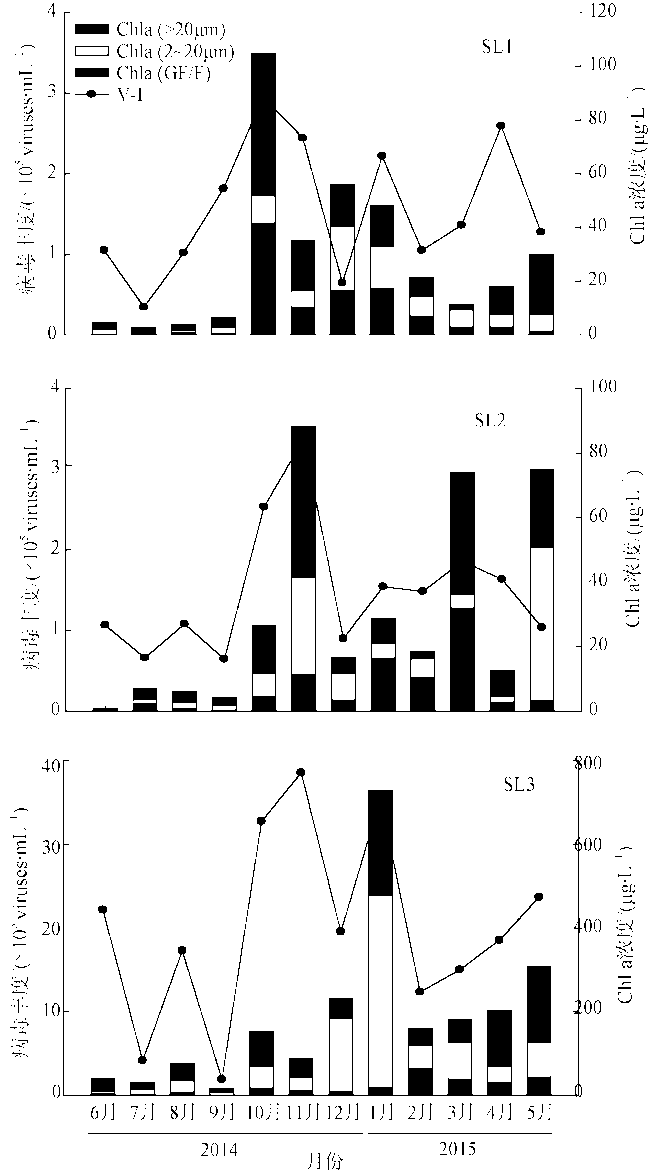
Fig. 8 Annual variation of V-I and Chl a at the three stations图8 三个站位的V-I病毒丰度与Chl a年度变化 |
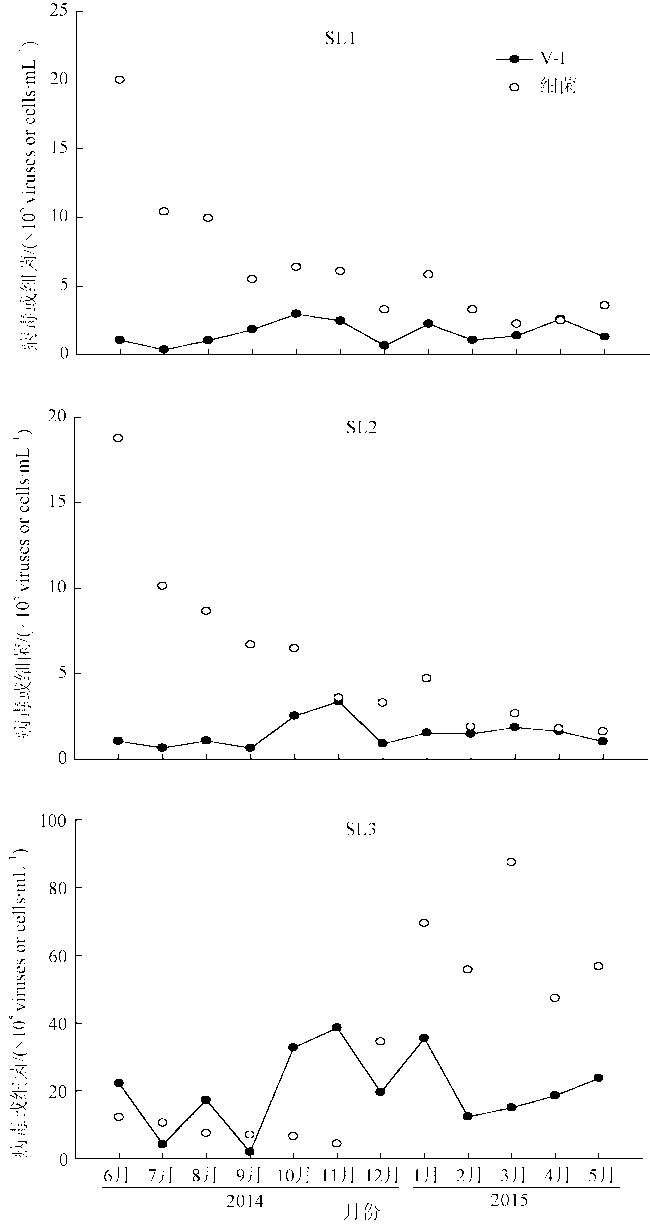
Fig. 9 Annual variation of V-I and bacterioplankton abundance at the three stations图9 三个站位SL1、SL2、SL3的V-I与浮游细菌丰度年度变化 |
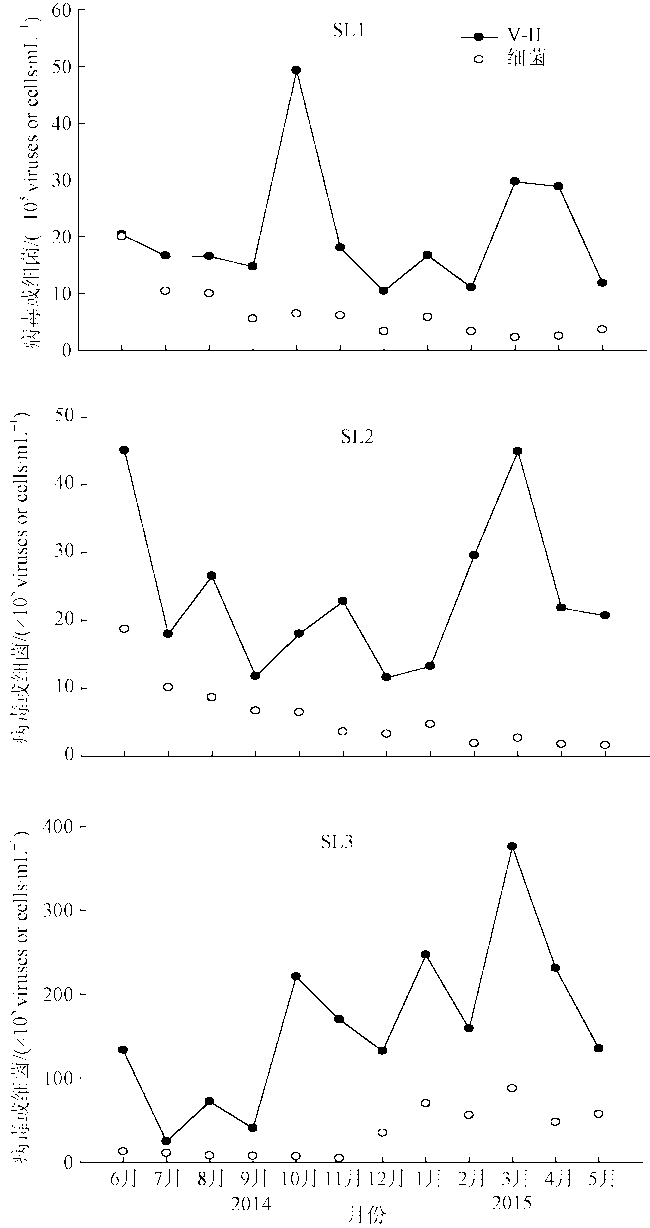
Fig. 10 Annual variation of V-Ⅱ and bacterioplankton abundance at the three stations图10 三个站位SL1、SL2、SL3的V-Ⅱ与浮游细菌丰度年度变化 |
Fig. 11 Annual variation of V-Ⅱ and Chl a at the three stations图11 三个站位SL1(左图)、SL2(中图)、SL3(右图)V-Ⅱ与Chl a年度变化 |
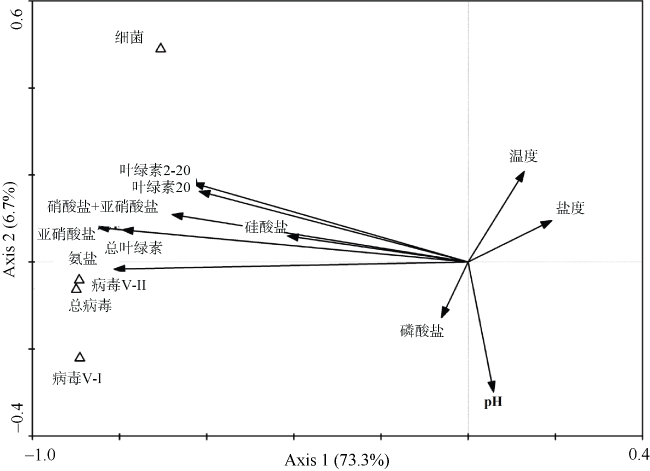
Fig. 12 RDA results of virioplankton, bacterioplankton and environmental parameters of the overall level of the surveyed stations图12 调查海域浮游病毒、浮游细菌与环境参数间的RDA分析结果 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
国家海洋局海口海洋环境监测中心站, 2005. 三亚湾海滩“泥化”和海岸侵蚀原因及治理对策研究报告[R]. 海口: 国家海洋局海口海洋环境监测中心站.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
WANG JIAN, WANG JI.1984The method for the determination of chlorophyll and phaeopigments in phytoplankton[J]. Wuhan Botanical Research, 2(2): 321-328 (in Chinese with English abstract).
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}