
Journal of Tropical Oceanography >
Effects of thermal discharge and nutrients input on size structure of phytoplankton in Daya Bay
Received date: 2017-07-25
Request revised date: 2017-09-26
Online published: 2018-05-03
Supported by
National Key Research and Development Program of China (2016YFC0502805)
National Basic Research Program (2015CB452904)
General Programs of the National Natural Science Foundation of China (41276161)
Copyright
Marine ecological effects induced by thermal discharge of nuclear plants in Daya Bay are attracting increasingly attention. Based on in situ data and laboratory experiments, distribution characteristics of phytoplankton size structure within the thermal-discharge-affected area and their responses to warming and nutrients input were explored. The results indicated that phytoplankton growth could be promoted by moderate warming but inhibited in extremely high temperature (36ºC). Chl-a concentrations near the outfall of thermal discharges were obviously low in both summer and winter. There were seasonal discrepancies between two important environmental factors, i.e., warming and nutrient enrichment, on their contributions to phytoplankton growth based on simulation results. Nutrients inputs showed more obvious impacts on phytoplankton than temperature in summer, and vice versa in winter. The increasing proportion of small-size phytoplankton (< 20μm) might be induced by warming and/or nutrient enrichment in Daya Bay, which confirmed the miniaturization trend within the plankton community and indicated its potential impacts on energy flow and material cycle, ecosystem stability, and production of marine fisheries in the bay.
Key words: phytoplankton; size structure; thermal discharge; nutrients; Daya Bay
XIE Fuwu , LIU Huaxue , HUANG Honghui , SONG Xingyu . Effects of thermal discharge and nutrients input on size structure of phytoplankton in Daya Bay[J]. Journal of Tropical Oceanography, 2018 , 37(3) : 55 -64 . DOI: 10.11978/2017083
Tab. 1 Sea surface temperature sampling stations表1 现场采样站位表层水温 |
| S1/W1 | S2/W2 | S3/W3 | S4/W4 | S5/W5 | S6/W6 | S7/W7 | |
|---|---|---|---|---|---|---|---|
| 夏季/℃ | 36.0 | 34.0 | 32.0 | 30.0 | 28.8 | — | — |
| 冬季/℃ | 28.3 | 26.2 | 24.0 | 22.0 | 21.0 | 20.0 | 19.0 |
注: S为夏季采样站位, W为冬季采样站位。 |
Tab. 2 Design of lab experiment during summer and winter表2 夏冬两季室内模拟实验设计 |
| 单因素模拟实验(M1) | 交互模拟实验(M2) | |
|---|---|---|
| 夏季 | 设置4个温度(36℃、32℃、28℃、26℃) | 设置3个温度梯度(36℃、32℃、28℃)和3个营养盐加富梯度 |
| 冬季 | 设置4个温度(28℃、24℃、20℃、18℃) | 设置3个温度梯度(28℃、24℃、20℃)和3个营养盐加富梯度 |
注: 营养盐加富梯度(低浓度组: 不添加营养盐; 中浓度组: 终浓度为10μmol·L-1的硝酸盐及0.6μmol·L-1的磷酸盐; 高浓度组: 终浓度为100μmol·L-1的硝酸盐及6μmol·L-1的磷酸盐。添加营养盐的选择依据朱艾嘉 等(2008)。 |
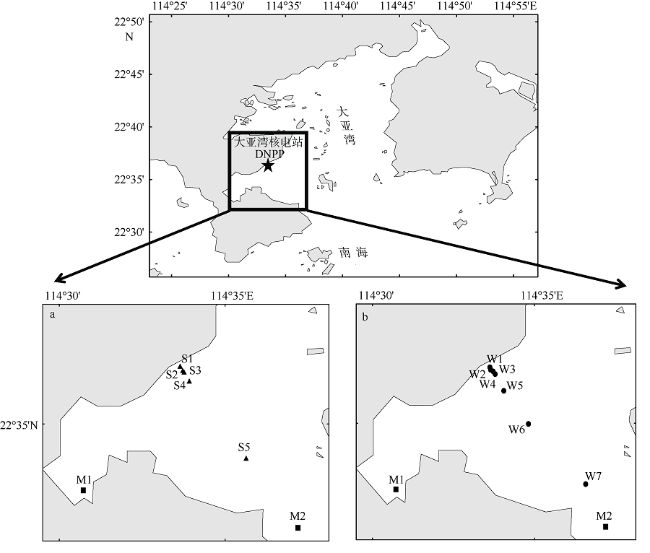
Fig. 1 Maps of sampling stations in Daya Bay: a) summer(sampling dates: 2015-08-17), and b) winter (sampling dates: 2015-12-21).▲: in situ sampling stations in summer;●: in situ sampling stations in winter; ■: sampling stations of simulated experiment图1 现场温度调查和模拟实验采样站位分布图 |
Tab. 3 Environmental parameters of surface water in Daya Bay during summer and winter表3 夏冬两季大亚湾表层代表性站位环境参数 |
| 季节 | 站位 | 温度/℃ | 盐度/‰ | pH | DIP /(μmol·L-1) | DIN /(μmol·L-1) | DSi /(μmol·L-1) | DIN/DIP | DSi/DIN | DSi/DIP |
|---|---|---|---|---|---|---|---|---|---|---|
| 夏季 | S1 | 36.0 | 31.78 | 8.07 | 0.17 | 4.98 | 13.38 | 29.29 | 2.69 | 78.71 |
| S2 | 34.0 | 31.92 | 8.11 | 0.23 | 7.11 | 15.35 | 30.91 | 2.16 | 66.74 | |
| S4 | 30.0 | 31.96 | 8.13 | 0.18 | 7.00 | 18.56 | 38.89 | 2.65 | 103.11 | |
| M1 | 28.2 | 32.26 | 8.25 | 0.33 | 16.61 | 16.28 | 50.33 | 0.98 | 49.33 | |
| M2 | 28.8 | 32.07 | 8.22 | 0.20 | 5.09 | 10.46 | 25.45 | 2.06 | 52.30 | |
| 冬季 | W1 | 28.3 | 33.86 | 8.14 | 0.26 | 11.71 | 16.78 | 45.04 | 1.43 | 64.54 |
| W2 | 26.2 | 33.79 | 8.17 | 0.18 | 13.13 | 11.56 | 72.94 | 0.88 | 64.22 | |
| W6 | 20.0 | 34.02 | 8.25 | 0.27 | 12.35 | 20.28 | 45.74 | 1.64 | 75.11 | |
| M1 | 18.4 | 33.61 | 8.15 | 0.12 | 25.26 | 10.68 | 210.50 | 0.42 | 89.00 | |
| M2 | 19.2 | 33.92 | 8.22 | 0.17 | 11.13 | 13.38 | 65.47 | 1.20 | 78.71 |
注: DIN为NO3-_ N、NO2-_ N和NH4+_ N浓度之和。 |
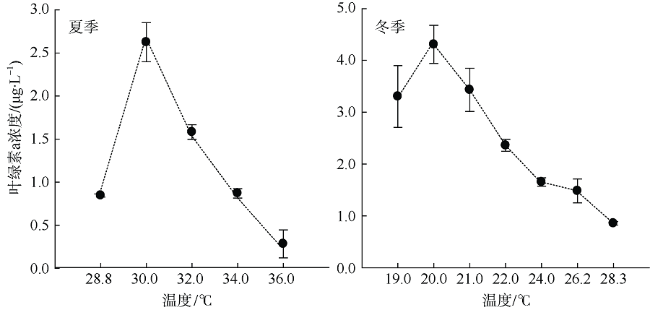
Fig. 2 Chl-a concentrations under difference in situ temperature gradients. Error bars are given图2 不同现场温度梯度叶绿素a含量 |
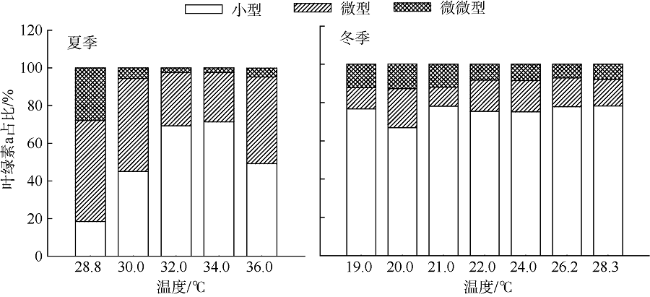
Fig. 3 Size-fractioned Chl-a under different in situ temperature gradients图3 不同现场温度梯度叶绿素a粒级结构 |
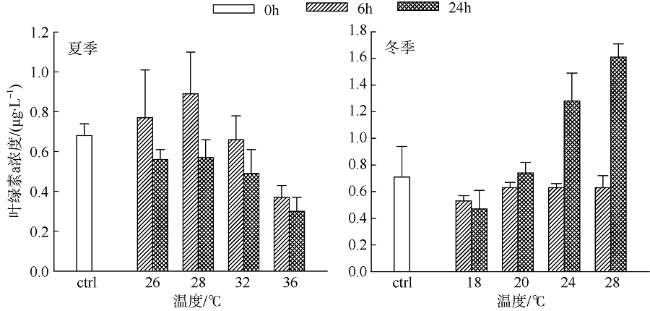
Fig. 4 Chl-a concentrations from simulated experiments. Error bars are given图4 模拟温度实验叶绿素a含量 |
Tab. 4 Distributions of size-fractionated Chl-a of simulated experiment in summer表4 夏季温度梯度实验浮游植物叶绿素a粒级结构分布 |
| 温度/℃ | 时间/h | 粒级 | ||
|---|---|---|---|---|
| 小型浮游 植物/% | 微型浮游 植物/% | 微微型 浮游植物/% | ||
| 28 | 0 | 13.7 | 63.6 | 22.7 |
| 26 | 6 | 17.7 | 59.0 | 23.3 |
| 24 | 21.2 | 63.3 | 15.5 | |
| 28 | 6 | 20.9 | 56.4 | 22.8 |
| 24 | 23.9 | 59.7 | 16.4 | |
| 32 | 6 | 20.8 | 64.0 | 15.2 |
| 24 | 27.1 | 59.2 | 13.7 | |
| 36 | 6 | 16.7 | 56.4 | 27.0 |
| 24 | 16.5 | 62.5 | 21.0 | |
Tab. 5 Distributions of size-fractionated Chl-a of simulated experiment in winter表5 冬季温度梯度实验浮游植物叶绿素a粒级结构分布 |
| 温度/℃ | 时间/h | 粒级 | ||
|---|---|---|---|---|
| 小型浮游 植物/% | 微型浮游 植物/% | 微微型 浮游植物/% | ||
| 20 | 0 | 9.3 | 19.0 | 71.7 |
| 18 | 6 | 9.9 | 34.7 | 55.4 |
| 24 | 19.5 | 33.0 | 47.5 | |
| 20 | 6 | 18.1 | 23.5 | 58.4 |
| 24 | 18.9 | 25.1 | 56.0 | |
| 24 | 6 | 13.3 | 30.7 | 56.0 |
| 24 | 17.2 | 27.3 | 55.5 | |
| 28 | 6 | 13.1 | 28.3 | 58.6 |
| 24 | 14.8 | 26.3 | 58.9 | |
Tab. 6 Chl-a concentrations of two-factors simulated experiment in winter表6 夏季双因素交叉模拟实验叶绿素a浓度 |
| 温度/℃ | 低营养/(μg·L-1) | 中营养/(μg·L-1) | 高营养/(μg·L-1) | 平均值/(μg·L-1) |
|---|---|---|---|---|
| 28 | 0.63±0.07 | 1.99±0.53 | 3.36±0.24 | 1.99±0.28 a |
| 32 | 0.48±0.08 | 1.88±0.77 | 2.75±0.78 | 1.70±0.54a |
| 36 | 0.66±0.17 | 2.24±1.05 | 2.42±1.86 | 1.77±1.02 a |
| 平均值 | 0.59±0.11a | 2.04±0.78 b | 2.84±0.96 c | — |
注: 叶绿素a初始浓度(现场海水温度28℃): 1.53±0.10μg·L-1。±后数值为标准差。标注字母不同的实验组之间存在显著性差异(p<0.05), 标注相同字母的实验组之间差异性不显著。 |
Tab. 7 Chl-a concentrations of two-factors simulated experiment in winter表7 冬季双因素交叉模拟实验叶绿素a浓度 |
| 温度/℃ | 低营养/(μg·L-1) | 中营养/(μg·L-1) | 高营养 /(μg·L-1) | 平均值/(μg·L-1) |
|---|---|---|---|---|
| 20 | 0.54±0.11 | 0.58±0.17 | 0.53±0.02 | 0.55±0.10 a |
| 24 | 1.10±0.17 | 1.08±0.07 | 1.38±0.12 | 1.19±0.12 b |
| 28 | 0.78±0.11 | 0.99±0.14 | 0.97±0.07 | 0.91±0.11 c |
| 平均值 | 0.81±0.13 a | 0.88±0.13 a | 0.96±0.07 a | — |
注: 叶绿素a初始浓度(现场海水温度20℃): 0.75±0.06μg·L-1。±后数值为标准差。标注字母不同的实验组之间存在显著性差异(p<0.05), 标注相同字母的实验组之间差异性不显著。 |
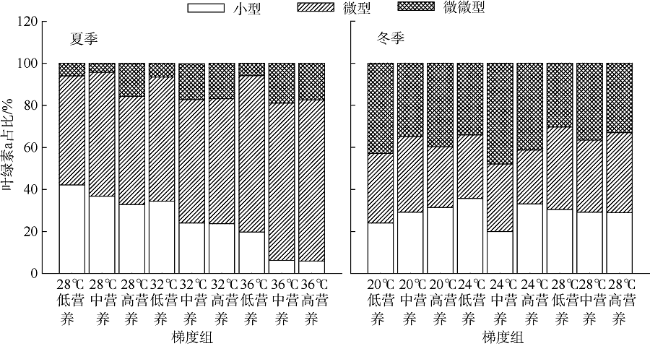
Fig. 5 Size-fractioned Chl-a from two-factor simulation experiments图5 双因素实验浮游植物叶绿素a粒级结构 |
Tab. 8 Pearson correlation coefficients between environmental factors and size-fractionated Chl-a表8 不同粒级叶绿素a与温度的Pearson相关系数 |
| 粒级 | 现场温度 梯度实验 | 单因子 模拟实验 | 双因子 模拟实验 | |||
|---|---|---|---|---|---|---|
| 夏季 | 冬季 | 夏季 | 冬季 | 夏季 | 冬季 | |
| 小型 浮游植物 | -0.278 | -0.931** | -0.772 | 0.943 | -0.657 | 0.427 |
| 微型 浮游植物 | -0.678 | -0.656 | -0.916 | 0.996** | 0.207 | 0.694* |
| 微微型 浮游植物 | -0.907* | -0.864* | -0.933 | 0.983* | 0.209 | 0.190 |
注: ** p<0.01, * p<0.05。 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. GB/T 12763.4-2007 海洋调查规范第4部分: 海水化学要素调查[S]. 北京: 中国标准出版社: 13-23.
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}