
Journal of Tropical Oceanography >
The ecological effect of marine microplastics as a biological vector
Received date: 2017-10-18
Request revised date: 2018-04-07
Online published: 2018-07-16
Supported by
National Key Research and Development Program of China (91328203);Young Scientists Fund of the National Natural Science Foundation of China (41406133);Natural Science Foundation of Guangdong Province (2015A030313071)
Copyright
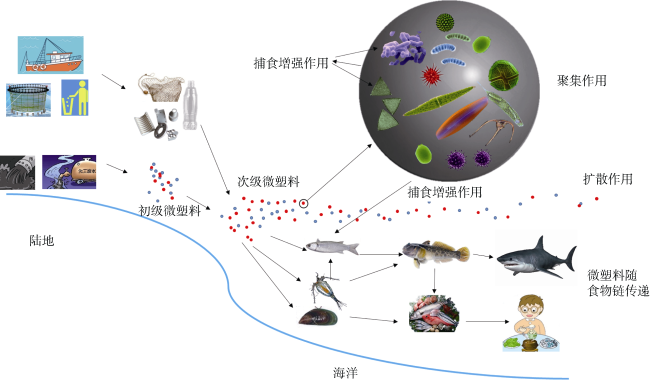
With the increase of marine plastic waste, marine microplastics have attracted more and more attention as a new type of marine pollution. At present, studies of the source, distribution and analytical approach of microplastics are well recognized. Most research focuses on plastic ingestion by marine organisms, and on the effects of microplastics adsorbing and releasing toxic chemicals. However, the role of microplastics as a vector for pelagic microorganism, phytoplankton and micozooplankton is poorly understood. In this review, we address three functions of microplastics as a biological vector. 1) Aggregation, it is easy for biofilms to form a microbial community on the surface of microplastics, which provide the means for horizontal transfer of organisms and genes, and may lead to the transformation or transduction of pathogenic gene, antibiotic resistance gene (ARGs), called genetic exchange. 2) Dispersal, the spread of harmful algal species, pathogenic bacteria and drug-resistant bacteria as well as other microorganisms may happen when the microplastics are freshly from the waste water flow, which potentially poses a risk of invasion when arriving in a new habitat with favorable condition. 3) A feeding enhancement concept, we propose this concept because a piece of microplastics is full of the attached biofilms, nutrients and organisms, which may attract large grazers or predators and can also improve their predation efficiency. As a result, these grazers/predators may seek pieces of microplastic as foods, which may cause more serious toxicological effects. The review focuses on the ecological effects of “microplastic + biology”.
Key words: microplastics; vector; genetic exchange; pathogenic bacteria; feeding enhancement
HE Lei , HUANG Fangjuan , YIN Kedong . The ecological effect of marine microplastics as a biological vector[J]. Journal of Tropical Oceanography, 2018 , 37(4) : 1 -8 . DOI: 10.11978/2017112
Fig. 1 The pathways of marine microplastics as a vector of ecological effects图1 海洋微塑料作为载体的生态效应模式图 |
The authors have declared that no competing interests exist.
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
BRETELER W C M K,
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
| 38 |
|
| 39 |
|
| 40 |
|
| 41 |
|
| 42 |
|
| 43 |
|
| 44 |
|
| 45 |
|
| 46 |
|
| 47 |
|
| 48 |
|
| 49 |
|
| 50 |
|
| 51 |
|
| 52 |
|
| 53 |
|
| 54 |
|
| 55 |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}