
Journal of Tropical Oceanography >
Grazing of three common protozoan on brown tide alga Aureococcus anophagefferens
Received date: 2017-12-13
Online published: 2019-05-06
Supported by
Natural Science Foundation of Guangdong Province (2017A030313217);Fundamental Research Funds for the Central Universities (21617450);National Marine Public Welfare Industry Research Project (201305003)
Copyright
With the increase of marine plastic waste, marine microplastics have attracted more and more attention as a new type of marine pollution. At present, studies of the source, distribution and analytical approach of microplastics are well recognized. Most research focuses on plastic ingestion by marine organisms, and on the effects of microplastics adsorbing and releasing toxic chemicals. However, the role of microplastics as a vector for pelagic microorganism, phytoplankton and micozooplankton is poorly understood. In this review, we address three functions of microplastics as a biological vector. 1) Aggregation, it is easy for biofilms to form a microbial community on the surface of microplastics, which provide the means for horizontal transfer of organisms and genes, and may lead to the transformation or transduction of pathogenic gene, antibiotic resistance gene (ARGs), called genetic exchange. 2) Dispersal, the spread of harmful algal species, pathogenic bacteria and drug-resistant bacteria as well as other microorganisms may happen when the microplastics are freshly from the waste water flow, which potentially poses a risk of invasion when arriving in a new habitat with favorable condition. 3) A feeding enhancement concept, we propose this concept because a piece of microplastics is full of the attached biofilms, nutrients and organisms, which may attract large grazers or predators and can also improve their predation efficiency. As a result, these grazers/predators may seek pieces of microplastic as foods, which may cause more serious toxicological effects. The review focuses on the ecological effects of “microplastic + biology”.
Key words: brown tide; A. anophagefferens; protozoan; grazing; EPS
CHEN Yao , YANG Xilu , HE Xuejia . Grazing of three common protozoan on brown tide alga Aureococcus anophagefferens[J]. Journal of Tropical Oceanography, 2018 , 37(6) : 120 -132 . DOI: 10.11978/2017130
Tab. 1 The food concentrations for growth and feeding experiments表1 生长和摄食实验的饵料浓度 |
| 饵料浓度 /(mg C·L-1) | 抑食金球藻细胞密度 /(×105cells·mL-1) | 球等鞭金藻细胞密度 /(×104cells·mL-1) |
|---|---|---|
| 0.2 | 1.38 | 2.16 |
| 0.3 | 2.07 | 3.24 |
| 0.5 | 3.44 | 5.40 |
| 1.0 | 6.88 | 10.8 |
| 1.5 | 10.3 | 16.2 |
| 3.0 | 20.6 | 32.4 |
Tab. 2 The experimental setup of three protozoan grazing on A. anophagefferens and I. galbana in mono-algal diet or in mixed diet表2 海洋尖尾藻、海洋尾丝虫、扇形游仆虫对抑食金球藻、球等鞭金藻的单种和混合饵料摄食实验 |
| 实验 | 设置 | 摄食者 | 处理 | 饵料浓度/(mg C·L-1) |
|---|---|---|---|---|
| 单种饵料生长 | 1 | 海洋尖尾藻 | 100%抑食金球藻 | 0.2、0.3、0.5、1.0、1.5、3.0 |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 | ||||
| 2 | 海洋尖尾藻 | 100%球等鞭金藻 | 0.2、0.3、0.5、1.0、1.5、3.0 | |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 | ||||
| 单种饵料摄食 | 3 | 海洋尖尾藻 | 100%抑食金球藻 | 0.2、0.3、0.5、1.0、1.5、3.0 |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 | ||||
| 4 | 海洋尖尾藻 | 100%球等鞭金藻 | 0.2、0.3、0.5、1.0、1.5、3.0 | |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 | ||||
| 混合饵料选择性摄食 | 5 | 海洋尖尾藻 | 抑食金球藻(指数期)∶球等鞭金藻为 80∶20、50∶50、20∶80 | 1.5 |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 | ||||
| 6 | 海洋尖尾藻 | 抑食金球藻(稳定期)∶球等鞭金藻为 80∶20、50∶50、20∶80 | 1.5 | |
| 海洋尾丝虫 | ||||
| 扇形游仆虫 |
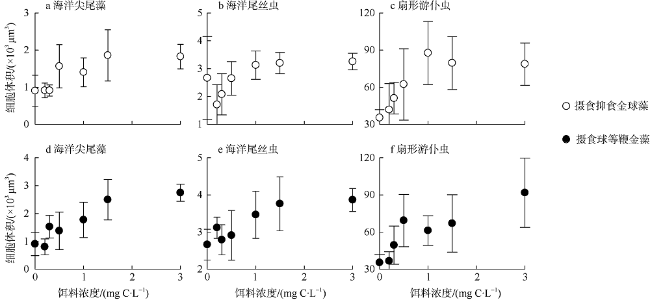
Fig. 1 Variation in the volumes of three protozoan grazing on A. anophagefferens or I. galbana at different food levels图1 三种原生动物摄食不同密度的抑食金球藻(a—c)和球等鞭金藻(d—f)的细胞体积变化 |
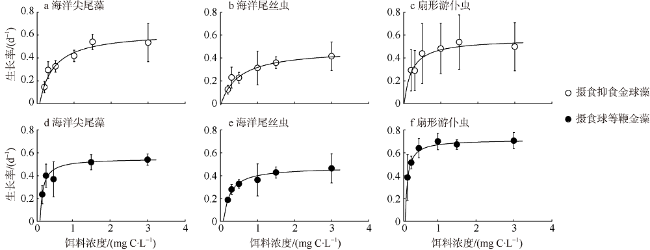
Fig. 2 Specific growth rates of three protozoan feeding on A. anophagefferens and I. galbana at different food levels图2 三种原生动物摄食不同细胞密度抑食金球藻(a—c)和球等鞭金藻(d—f)的生长率 |
Tab. 3 Analysis of functional response of physiological variables, growth rate, and ingestion rate to prey concentration in three protozoa feeding on A. anophagefferens and I. galbana, respectively表3 三种原生动物的生理参数以及生长率、摄食率对饵料浓度的响应的拟合回归分析 |
| 参数 | 摄食抑食金球藻 | 摄食球等鞭金藻 | ||||
|---|---|---|---|---|---|---|
| 海洋尖尾藻 | 海洋尾丝虫 | 扇形游仆虫 | 海洋尖尾藻 | 海洋尾丝虫 | 扇形游仆虫 | |
| μmax/(d-1) | 0.63±0.11 | 0.48±0.06 | 0.57±0.07 | 0.55±0.03 | 0.48±0.02 | 0.72±0.02 |
| KGR/(mg C·L-1) | 0.38±0.26 | 0.50±0.25 | 0.20±0.18 | 0.08±0.05 | 0.17±0.05 | 0.06±0.02 |
| r2 | 0.91 | 0.96 | 0.83 | 0.93 | 0.98 | 0.92 |
| Imax/(ng C·grazer-1·d-1) | 0.64±0.15 | 3.04±0.74 | 167.64±25.01 | 1.76±0.50 | 11.83±3.49 | 329.33±75.92 |
| KIR/(mg C·L-1) | 0.88±0.48 | 4.77±1.66 | 1.15±0.38 | 1.58±0.89 | 4.71±2.00 | 2.77±1.06 |
| r2 | 0.86 | 0.99 | 0.95 | 0.89 | 0.98 | 0.96 |
| GGEmax | 65.8%±13.0% | 35.2%±6.0% | 49.1%±14.6% | 43.3%±11.6% | 34.8%±5.4% | 76.5%±2.7% |
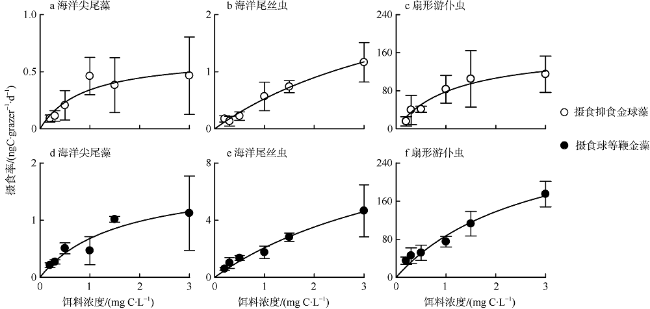
Fig. 3 Ingestion rates of three protozoan feeding on A. anophagefferens and I. galbana at different food levels图3 三种原生动物摄食不同细胞密度的抑食金球藻(a—c)和球等鞭金藻(d—f)的摄食率 |
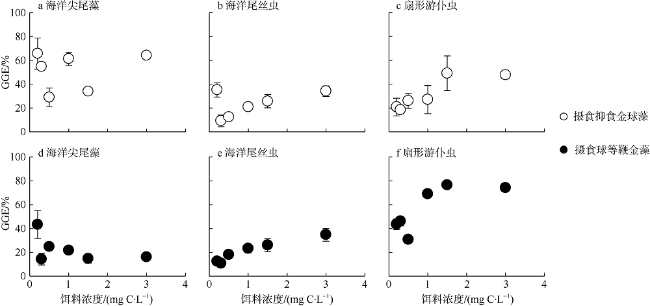
Fig. 4 Growth gross efficiencies of three protozoa feeding on A. anophagefferens and I. galbana at different food concentrations图4 三种原生动物分别摄食不同浓度抑食金球藻(a—c)和球等鞭金藻(d—f)时的毛生长率 |
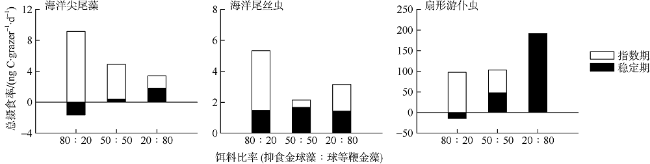
Fig. 5 Total ingestion rates of O. marina, U.marinum, and E. vannus on mixed diets图5 三种原生动物摄食混合饵料的总摄食率 |
Tab. 4 Selectivity index for O. marina, U. marinum, and E. vannus feeding on mixtures of cells of A. anophagefferens at different growth phases and I. galbana表4 海洋尖尾藻、海洋尾丝虫、扇形游仆虫对混合饵料中生长指数期和稳定期抑食金球藻的摄食选择性 |
| 摄食组 | 比例 | 饵料 | 抑食金球藻指数期 | 抑食金球藻稳定期 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FR | Ivlev's index | Jacob's index | 摄食行为 | FR | Ivlev's index | Jacob's index | 摄食行为 | |||||
| lgQ | D | lgQ | D | |||||||||
| 海洋尖尾藻 | 80∶20 | 抑食金球藻 | 1.136 | 0.064 | 0.396 | 0.427 | + | 0.997 | -0.003 | -0.007 | -0.008 | 0 |
| 球等鞭金藻 | 0.456 | -0.374 | -0.396 | -0.427 | - | 1.013 | 0.006 | 0.007 | 0.008 | 0 | ||
| 50∶50 | 抑食金球藻 | 1.655 | 0.247 | 0.681 | 0.655 | + | 0.104 | -0.812 | -1.263 | -0.896 | - | |
| 球等鞭金藻 | 0.345 | -0.487 | -0.681 | -0.655 | - | 1.896 | 0.309 | 1.263 | 0.896 | + | ||
| 20∶80 | 抑食金球藻 | 2.935 | 0.492 | 0.755 | 0.701 | + | 0.917 | -0.044 | -0.047 | -0.054 | 0 | |
| 球等鞭金藻 | 0.516 | -0.319 | -0.755 | -0.701 | - | 1.021 | 0.010 | 0.047 | 0.054 | 0 | ||
| 海洋尾丝虫 | 80∶20 | 抑食金球藻 | 1.061 | 0.030 | 0.148 | 0.169 | + | 0.916 | -0.044 | -0.164 | -0.187 | - |
| 球等鞭金藻 | 0.755 | -0.140 | -0.148 | -0.169 | - | 1.336 | 0.144 | 0.164 | 0.187 | + | ||
| 50∶50 | 抑食金球藻 | 1.125 | 0.059 | 0.109 | 0.125 | + | 0.599 | -0.251 | -0.369 | -0.401 | - | |
| 球等鞭金藻 | 0.875 | -0.066 | -0.109 | -0.125 | - | 1.401 | 0.167 | 0.369 | 0.401 | + | ||
| 20∶80 | 抑食金球藻 | 2.964 | 0.495 | 0.765 | 0.707 | + | 0.198 | -0.670 | -0.783 | -0.717 | - | |
| 球等鞭金藻 | 0.509 | -0.325 | -0.765 | -0.707 | - | 1.201 | 0.091 | 0.783 | 0.717 | + | ||
| 扇形游仆虫 | 80∶20 | 抑食金球藻 | 1.125 | 0.059 | 0.351 | 0.384 | + | 0.494 | -0.338 | -0.786 | -0.719 | - |
| 球等鞭金藻 | 0.501 | -0.332 | -0.351 | -0.384 | - | 3.023 | 0.503 | 0.786 | 0.719 | + | ||
| 50∶50 | 抑食金球藻 | 1.201 | 0.092 | 0.177 | 0.201 | + | 0.889 | -0.059 | -0.097 | -0.111 | 0 | |
| 球等鞭金藻 | 0.799 | -0.112 | -0.177 | -0.201 | - | 1.111 | 0.053 | 0.097 | 0.111 | 0 | ||
| 20∶80 | 抑食金球藻 | 1.575 | 0.223 | 0.265 | 0.296 | + | 0.973 | -0.014 | -0.015 | -0.017 | 0 | |
| 球等鞭金藻 | 0.856 | -0.077 | -0.265 | -0.296 | - | 1.007 | 0.003 | 0.015 | 0.017 | 0 | ||
注: 摄食行为中“-”表示选择性避食, “+”表示选择性摄食, “0”表示选择性较弱。各选择性指数的范围分别为, FR: 0~1 (-), 1~∞(+); Ivlev's index: -1~0(-), 0~1(+); Jacob's index: lgQ: -∞~0(-), 0~∞(+); D: -1~0(-), 0~1(+)。 |
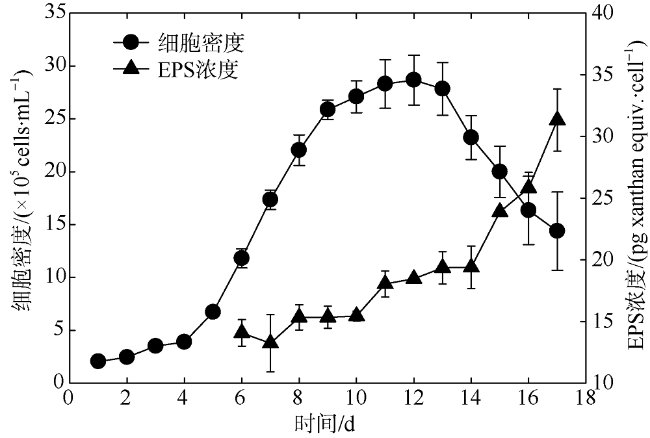
Fig. 6 EPS concentrations of A. anophagefferens culture at exponential growth phase and stationary growth phase, respectively图6 抑食金球藻各个生长时期细胞的EPS含量 |
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}