
Journal of Tropical Oceanography >
Energy accumulation and allocation of somatic and reproductive tissues in medium form of male Sthenoteuthisoualaniensis in the South China Sea
Copy editor: SUN Shu-jie
Received date: 2018-10-01
Request revised date: 2019-02-22
Online published: 2019-07-21
Supported by
National Natural Science Foundation of China((NSFC41276156);)
National High TechnologyResearch and Development Program (863 Program)((2012AA092303);)
Natural Science Foundation of Shanghai (16ZR1415400)((16ZR1415400);)
Shanghai Ocean University College Students' innovation training program((B1-5002-17-0001-2).)
Copyright
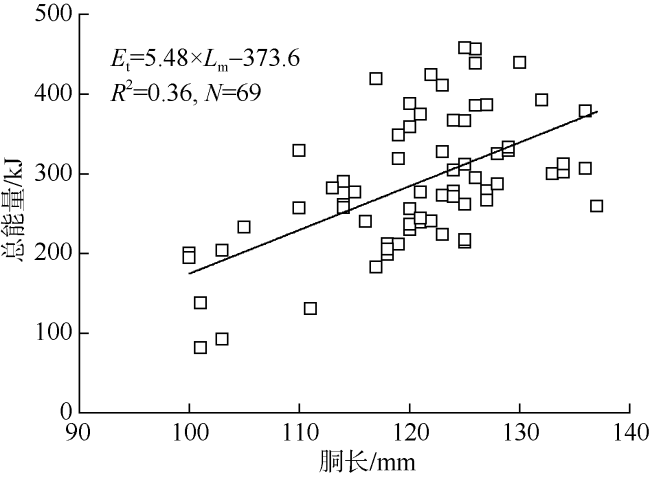
The energy accumulation and allocation of somatic and reproductive tissues in the medium form of male Sthenoteuthisoualaniensiswere were analyzed by using the technology of energy density determination combined with residual analysis. The results showed that the energy density of mantle, fins, arms, spermatophoric complex, and testis were 21.16±1.44, 21.98±1.51, 21.44±1.50, 20.83±1.70, and 21.41±1.70kJ·g -1, respectively. The energy density for tissue of mantle or spermatophoric complex varied significantly depending on sexual maturity stages, while there were no significant differences between sexual maturity stages for the tissue of fins, arms, or testis. The total energy accumulation for each individual was estimated at 299.31±90.81 kJ, and showed an increasing trend with body size growth. With regards to relative energy accumulation, the somatic tissues, including mantle, fins and arms, accounted for 95.31%~98.04% of the total energy accumulation, but decreased with maturation. The relative energy accumulation in reproductive tissues, being testis and spermatophoric complex, accounted for 1.96%~4.96% of the total energy accumulation, and increased with maturation, in which the maximum value was reached at stage V. The analysis of residuals of relationship between tissue absolute energy accumulation and mantle length, using Model II regression, showed a significantly positive correlation between somatic and gonadal tissues. This evidence suggests that there was no energy remobilization of somatic tissues to fuel reproduction during sexual maturation, and the mode of reproductive investment is more likely to be income breeding in the medium form of male S.oualaniensis. In this study, we preliminary established the progress of energy accumulation and its allocation in soma and reproductive organs for medium form of male S.oualaniensisin the South China Sea. These findings should assist future research on its reproductive strategy, as well as sustainable utilizationof this kind of resource in the area.
ZHU Kai , ZHANG Lichuan , CHEN Xinjun , YAO Jixiang , HAN Fei , LIN Dongming . Energy accumulation and allocation of somatic and reproductive tissues in medium form of male Sthenoteuthisoualaniensis in the South China Sea[J]. Journal of Tropical Oceanography, 2019 , 38(4) : 41 -51 . DOI: 10.11978/2018100
表1 鸢乌贼中型群雄性个体能量积累及其分配研究的样本组成Tab. 1 Specimens of the medium form of male S.oualaniensisused for the analysis of energy accumulation and allocation |
| 性腺成熟度 | 样本数 | 胴长/mm | 体重/g |
|---|---|---|---|
| Ⅰ | 11 | 100~121 (107.27±7.62) | 26.1~61.0 (41.51±9.23) |
| Ⅱ | 9 | 118~125 (121.56±2.71) | 44.2~75.0 (59.81±9.68) |
| Ⅲ | 12 | 113~136 (122.67±6.82) | 55.6~115.6 (72.97±17.50) |
| Ⅳ | 22 | 110~136 (125.68±5.57) | 53.0~105.7 (82.49±12.75) |
| Ⅴ | 15 | 110~137 (122.20±7.36) | 32.6~100.7 (68.22±18.24) |
| 总计 | 69 | 110~137 (120.63±10.57) | 26.1~100.7 (70.31±10.57) |
注: 括号内为平均值±SD。 |
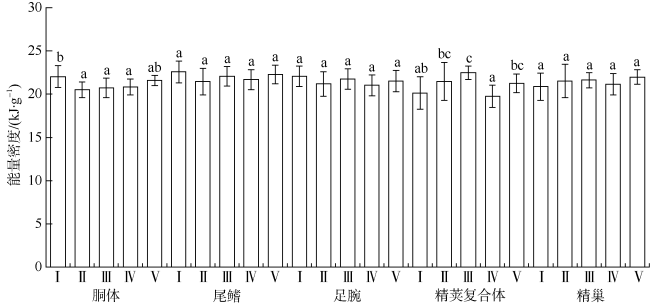
图2 不同性腺成熟度鸢乌贼中型群雄性个体的组织能量密度各组平均值中标示的不同字母表示Tukey HSD检验差异显著(P<0.05), 下同。样本数69 Fig. 2 The tissue energy density in the medium form of male S. oualaniensis according to different maturity. Mean values with different scripts are significantly different (P<0.05) as determined using Tukey HSD test; the same below. Sample number is 69 |
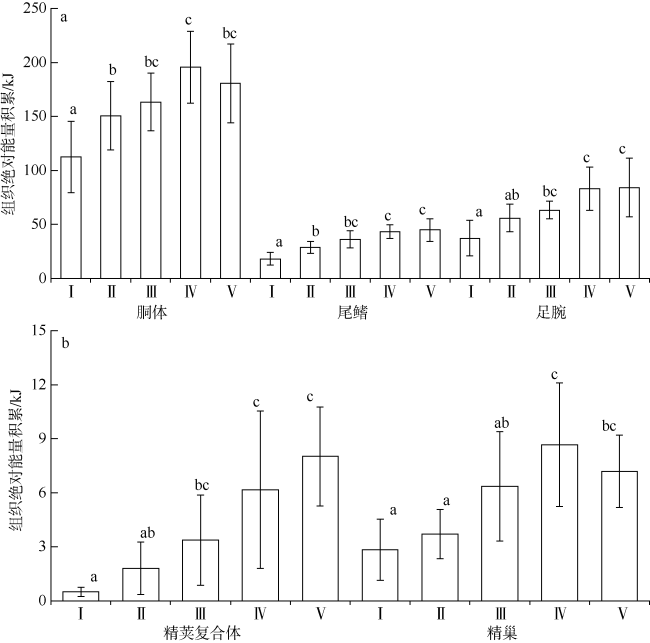
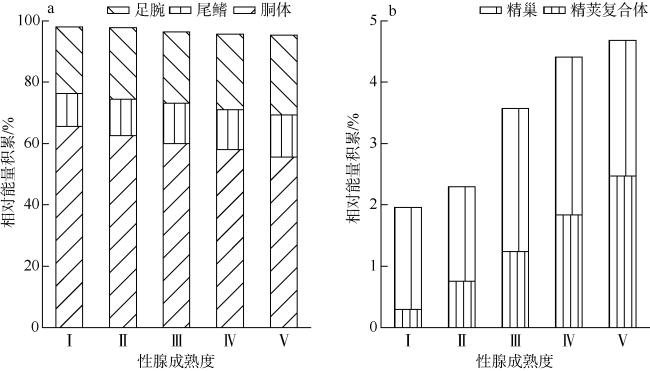
图3 不同性腺成熟度鸢乌贼中型群雄性个体各组织绝对能量积累分布a. 胴体、尾鳍和足腕组织绝对能量积累; b. 精荚复合体和精巢组织绝对能量积累。样本量为69 Fig. 3 The absolute energy accumulation in medium form of male S.oualaniensis according to different maturity. a) Absolute energy accumulation for mantle, fins and arms; b) absolute energy accumulation for spermatophoric complex and testis. Sample number is 69 |
图5 不同性腺成熟度鸢乌贼中型群雄性个体肌肉组织和性腺组织的相对能量积累a. 肌肉组织相对能量积累; b. 性腺组织相对能量积累。样本量为69 Fig. 5 Relative energy accumulation in soma and reproductive tissues with maturity in the medium form of male S.oualaniensis. a) Relative energy accumulation in somatic tissues; b) relative energy accumulation in reproductive tissues. Sample number is 69 |
表2 鸢乌贼中型群雄性个体胴体、尾鳍、足腕、性腺等组织绝对能量积累与胴长的Model Ⅱ回归参数表Tab. 2 Model Ⅱ regression statistics between absolute energy accumulation of mantle (Eama), fins (Eafi), arms (Eaar), and gonad (Eago) and mantle length (Lm) for the medium form of maleS.oualaniensis |
| 组织能量-胴长 | 截距 | 截距95%置信区间 | 斜率 | 斜率95%置信区间 | R2 | 尾数 |
|---|---|---|---|---|---|---|
| Eama-Lm | -131.31 | -256.89 ~ -5.73 | 2.47 | 1.44~3.51 | 0.25 | 69 |
| Eafi-Lm | -84.22 | -112.43 ~ -56.00 | 1.00 | 0.77~1.23 | 0.52 | 69 |
| Eaar-Lm | -91.04 | -167.84 ~ -14.23 | 1.32 | 0.69~1.96 | 0.21 | 69 |
| Eago-Lm | -43.54 | -60.98 ~ -26.09 | 0.45 | 0.31~0.59 | 0.37 | 69 |
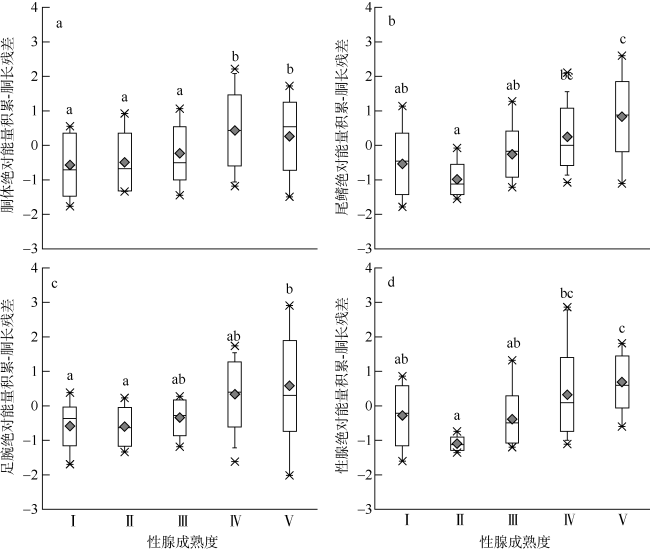
图6 不同性腺成熟度鸢乌贼中型群雄性个体肌肉和性腺组织绝对能量积累-胴长残差分布a. 胴体绝对能量积累-胴长残差; b. 尾鳍绝对能量积累-胴长残差; c. 足腕绝对能量积累-胴长残差; d. 性腺绝对能量积累-胴长残差 Fig. 6 Distributions of residuals from the regressions of somatic and reproductive tissues absolute energy accumulation on mantle length in the medium form of male S.oualaniensis. a) Eama-Lm residuals; b) Eafi-Lm residuals; c) Eaar-Lm residuals; d) Eago-Lm residuals |
表3 鸢乌贼中型群雄性个体性腺组织绝对能量积累-胴长残差与肌肉组织绝对能量积累-胴长残差的Pearson相关性Tab. 3 Pearson correlation of residuals from reproductive tissues absolute energy accumulation-mantle length regression with somatic tissues absolute energy accumulation-mantle length residuals for medium form of male S.oualaniensis |
| 性腺成熟度 | 性腺vs.胴体 | 性腺vs.尾鳍 | 性腺vs.足腕 | |||
|---|---|---|---|---|---|---|
| Pearson相关性 | 显著性 | Pearson相关性 | 显著性 | Pearson相关性 | 显著性 | |
| 总体 | 0.39 | 0.001 | 0.67 | 0.009 | 0.67 | 0 |
| Ⅰ | 0.62 | 0.04 | 0.85 | 0.001 | 0.11 | 0.74 |
| Ⅱ | -0.52 | 0.15 | 0.15 | 0.70 | -0.76 | 0.02 |
| Ⅲ | 0.54 | 0.07 | 0.36 | 0.25 | 0.19 | 0.78 |
| Ⅳ | -0.01 | 0.961 | 0.33 | 0.14 | -0.30 | 0.17 |
| Ⅴ | 0.59 | 0.021 | 0.74 | 0.002 | 0.68 | 0.005 |
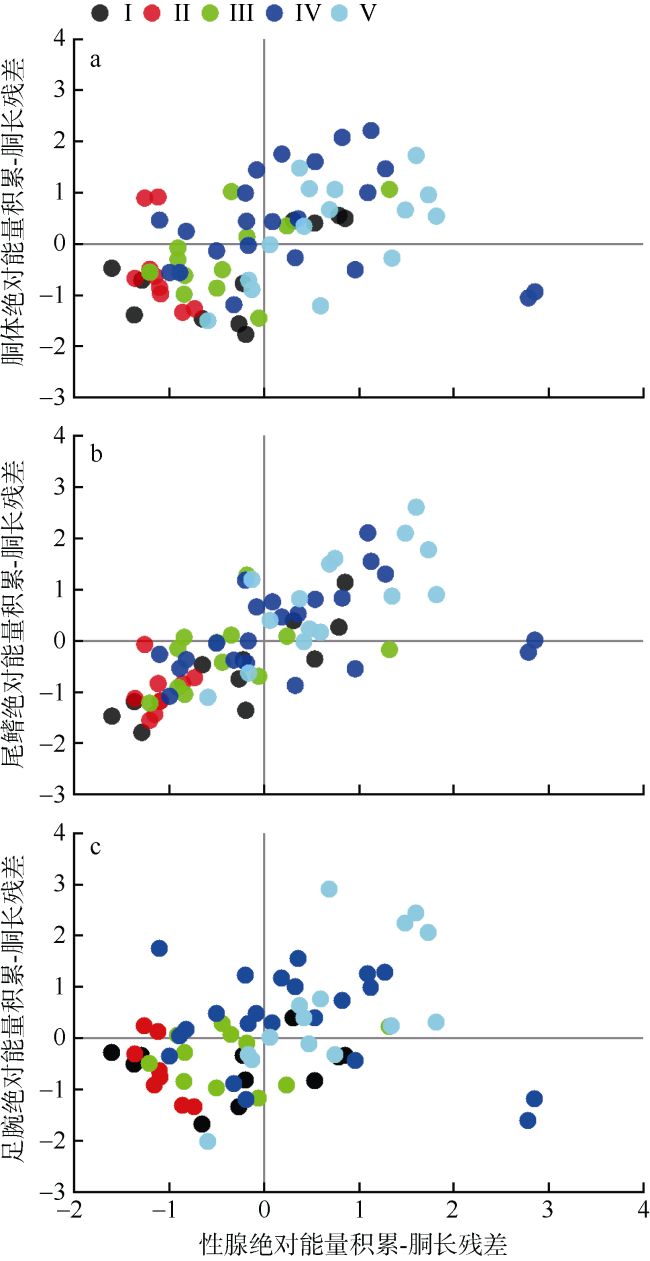
图7 鸢乌贼中型群雄性个体胴体、尾鳍、足腕、性腺组织绝对能量积累与胴长Model Ⅱ回归拟合的残差关系图a. 胴体绝对能量积累-胴长残差与性腺绝对能量积累-胴长残差; b. 尾鳍绝对能量积累-胴长残差与性腺绝对能量积累-胴长残差; c. 足腕绝对能量积累-胴长残差与性腺绝对能量积累-胴长残差 Fig. 7 Relationships between residuals calculated from absolute energy accumulation-mantle length Model Ⅱ regression equations for mantle, fins, arms, and gonad in the medium form of male S.oualaniensis. a) Eama-Lm residuals vs. Eago-Lm residuals; b) Eafi-Lm residuals vs. Eago-Lm residuals; c) Eaar-Lm residuals vs. Eago-Lm residuals |
| [1] |
陈新军, 刘必林, 王尧耕 , 2009. 世界头足类[M]. 北京: 海洋出版社: 312-313.
|
| [2] |
崔奕波 , 1989. 鱼类生物能量学的理论与方法[J]. 水生生物学报, 13(4):369-383.
|
| [3] |
戴宏杰, 陈道海 , 2014. 头足类营养研究进展[J]. 动物营养学报, 26(3):597-604.
|
| [4] |
范江涛, 冯雪, 邱永松 , 等, 2013. 南海鸢乌贼生物学研究进展[J]. 广东农业科学, 40(23):122-128.
|
| [5] |
龚玉艳, 詹凤娉, 杨玉滔 , 等, 2016. 南海鸢乌贼摄食习性的初步研究[J]. 南方水产科学, 12(4):80-87.
|
| [6] |
江艳娥, 陈作志, 林昭进 , 等, 2019. 南海海域鸢乌贼中型群与微型群渔业生物学比较[J]. 水产学报, 43(2):454-466.
|
| [7] |
林东明 , 2015. 阿根廷滑柔鱼的产卵策略研究[D]. 上海: 上海海洋大学: 1-100.
|
| [8] |
林东明, 陈新军, 魏嫣然 , 等, 2017. 阿根廷滑柔鱼雌性个体肌肉和性腺组织能量积累及其生殖投入[J]. 水产学报, 41(1):70-80.
|
| [9] |
刘必林 , 2006. 利用耳石微结构研究印度洋西北海域鸢乌贼的年龄和生长[D]. 上海: 上海水产大学.
|
| [10] |
颜云榕, 冯波, 卢伙胜 , 等, 2012. 南沙群岛北部海域鸢乌贼(Sthenoteuthis oualaniensis)夏季渔业生物学研究[J]. 海洋与湖沼, 43(6):1177-1186.
|
| [11] |
张俊, 陈国宝, 张鹏 , 等, 2014. 基于渔业声学和灯光罩网的南海中南部鸢乌贼资源评估[J]. 中国水产科学, 21(4):822-831.
|
| [12] |
张鹏, 晏磊, 杨炳忠 , 等, 2015. 春季南沙海域鸢乌贼种群结构特征的研究[J]. 南方水产科学, 11(5):11-19.
|
| [13] |
张宇美, 颜云榕, 卢伙胜 , 等, 2013. 西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究[J]. 广东海洋大学学报, 33(3):56-64.
|
| [14] |
张宇美 , 2014. 基于碳氮稳定同位素的南海鸢乌贼摄食生态与营养级研究[D]. 湛江: 广东海洋大学: 1-53.
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}