
Journal of Tropical Oceanography >
Micro-bioerosion in Porites corals in the northern South China Sea
Copy editor: LIN Qiang
Received date: 2019-01-09
Request revised date: 2019-02-26
Online published: 2019-11-26
Supported by
National Science Foundation of China(41476038)
National Science Foundation of China(41676049)
Youth Innovation Promotion Association CAS(2015284)
Natural Science Foundation of Guangdong Province(2018A030313142)
Copyright
The effect of microorganisms on coral skeletons is more concealed than that by macro-bioerosion, which is visible to the naked eye. The study of micro-bioerosion is rare, and there is no such research in China. In this study, micro-bioerosion research was carried out under scanning electron microscopy on the coral skeletons of the Porites samples collected from Weizhou Island, Daya Bay and Chenhang Island. The “Orange Bands” were formed by microorganisms in the coral skeletons of Weizhou Island and Daya Bay, while there were no similar bands in the samples of Chenhang Island. This phenomenon may be related to the eutrophication of local sea water. Scanning electron microscopy of these "Orange Bands" revealed that this was caused by an endolithic algae—Ostreobium quekettii, a common eroding microorganism in the tropical coral reef area. The structure of Porites skeleton eroded by Ostreobium quekettii is destroyed, the thickness is reduced, the porosity is increased from 1.54% to 6.76%, and the compressive strength is reduced from 27.1% to 51.2%. The microorganism makes the coral skeleton more "loose" and form a layer of "osteoporosis" on the entire surface of the coral. This microorganism acts as a “forerunner,” making corals more susceptible to macro-bioerosion, which in turn promote micro-bioerosion. The “positive feedback effect” explains the increased bioerosion of the coral skeleton in the eutrophic northern South China Sea.
ZHAO Kuan , ZHANG Ting , CHEN Tianran . Micro-bioerosion in Porites corals in the northern South China Sea[J]. Journal of Tropical Oceanography, 2019 , 38(6) : 74 -79 . DOI: 10.11978/2019006
图2 珊瑚切片以及“橘色条带”(微生物侵蚀)在扫描电镜下的特征a、b、c为涠洲岛采集样品及扫描电镜图像; d、e为大亚湾采集样品及扫描电镜图像; f、g为琛航岛采集样品及扫描电镜图像 Fig. 2 “Orange Bands” (micro-bioerosion) on coral slices and skeletal micro-structures revealed by scanning electron microscopy. a, b and c are samples and SEM images of Weizhou Island; d, e and f are samples and SEM images of Daya Bay; f and g are samples and SEM images of Chenhang Island |
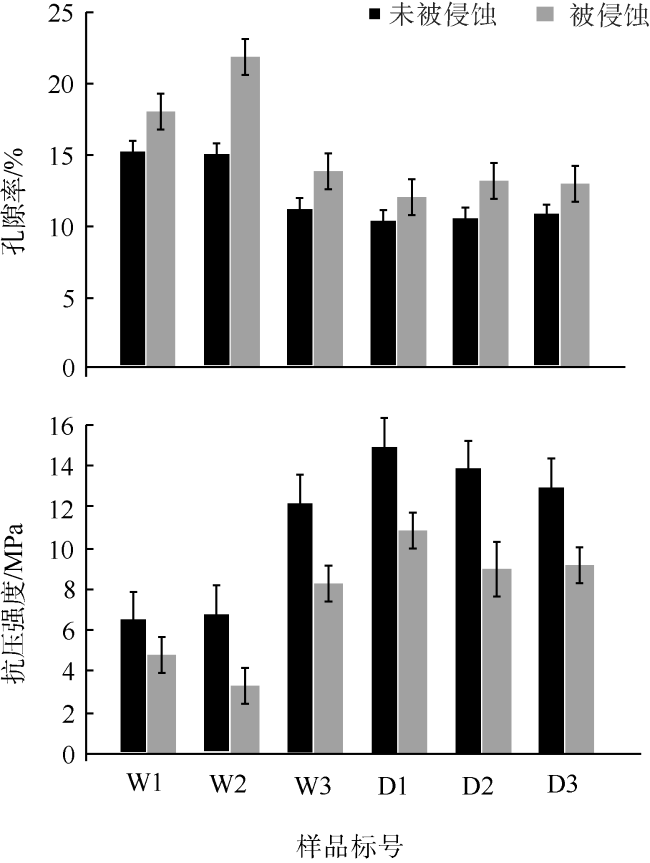
表1 被微生物侵蚀的珊瑚骨骼以及对照样品的物理参数变化Tab. 1 Differences in physical parameters between eroded and control coral skeletal samples |
| 样品 标号 | 孔隙率 /% | 干燥纵波波速/(m·s-1) | 抗压强度 /MPa | |||
|---|---|---|---|---|---|---|
| 未被侵蚀 | 被侵蚀 | 未被侵蚀 | 被侵蚀 | 未被侵蚀 | 被侵蚀 | |
| W1 | 15.24 | 17.98 | 3293.64 | 3062.83 | 6.65 | 4.84 |
| W2 | 15.02 | 21.78 | 3314.05 | 2792.21 | 6.84 | 3.33 |
| W3 | 11.21 | 13.66 | 3745.72 | 3448.53 | 12.39 | 8.23 |
| D1 | 10.30 | 11.84 | 3889.35 | 3659.76 | 15.10 | 11.01 |
| D2 | 10.60 | 13.01 | 3838.78 | 3518.99 | 14.08 | 9.07 |
| D3 | 10.91 | 12.95 | 3790.06 | 3525.73 | 13.17 | 9.15 |
| C1 | 9.43 | 4064.99 | 19.22 | |||
| C2 | 10.08 | 3929.05 | 15.94 | |||
| C3 | 10.06 | 3932.79 | 16.03 | |||
注: W为涠洲岛样品, D为大亚湾样品, C为琛航岛样品 |
| 1 |
陈刚, 赵美霞, 刘斌 , 等, 2016. 基于 Reef Check 调查的涠洲岛珊瑚礁生态状况评价[J]. 热带地理, 36(1):66-71.
|
| 2 |
陈天然, 余克服, 施祺 , 等, 2009. 大亚湾石珊瑚群落近25年的变化及其对2008年极端低温事件的响应[J]. 科学通报, 54(6):812-820.
|
| 3 |
陈天然, 郑兆勇, 莫少华 , 等, 2013. 涠洲岛滨珊瑚中的生物侵蚀及其环境指示意义[J]. 科学通报, 58:1574-1582.
|
| 4 |
黄晖, 马斌儒, 练健生 , 等, 2009. 广西涠洲岛海域珊瑚礁现状及其保护策略研究[J]. 热带地理, 29(4):307-312.
|
| 5 |
金昱昕, 陈天然, 孟庆山 , 等, 2017. 单轴抗压强度揭示南海珊瑚骨骼结构的差异[J]. 热带海洋学报, 36(2):33-39.
|
| 6 |
梁文, 黎广钊, 张春华 , 等, 2010. 20年来涠洲岛珊瑚礁物种多样性演变特征研究[J]. 海洋科学, 34(12):78.
|
| 7 |
梁鑫, 彭在清 , 2018. 广西涠洲岛珊瑚礁海域水质环境变化研究与评价[J]. 海洋开发与管理, 1:114-119.
|
| 8 |
聂永康, 陈丕茂, 余景 , 等, 2016. 大亚湾中央列岛海域水质分析[J]. 安徽农业科学, 44(16):57-60.
|
| 9 |
杨永康, 丁学武, 冯春燕 , 等, 2016. 西沙群岛珊瑚礁灰岩物理力学特性试验研究[J]. 广州大学学报, 15(5):78-82.
|
| 10 |
张颖, 杨清松, 张燕英 , 等, 2018. 造礁石珊瑚共附生固氮微生物的固氮活性[J]. 生态学杂志, 37(7):2122-2129.
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}