
Journal of Tropical Oceanography >
Seasonal succession of macroalgae community in Naozhou Island
Received date: 2019-03-09
Request revised date: 2019-08-06
Online published: 2020-01-09
Supported by
China Marine Special Nonprofit Scientific Research(201105008-5)
China Marine Special Nonprofit Scientific Research(201505027)
Copyright
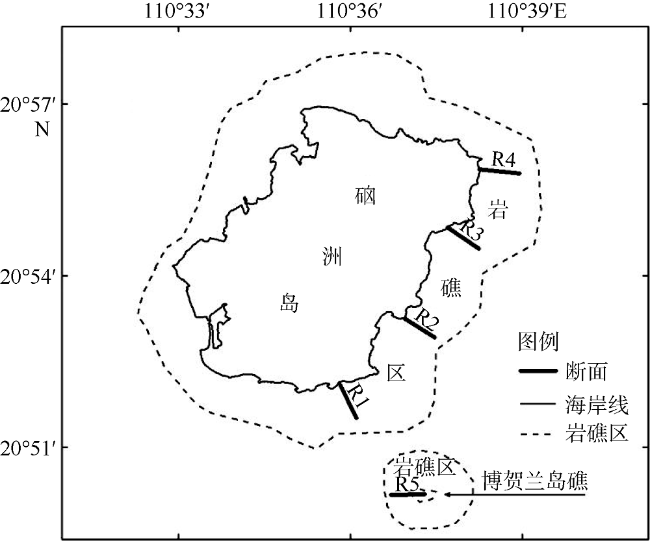
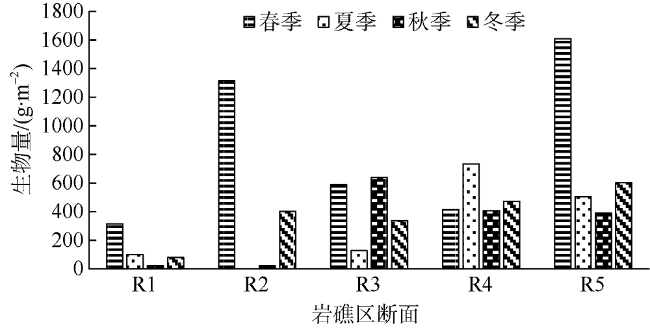
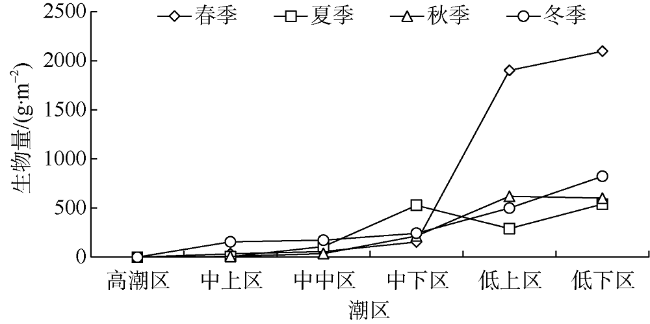
Seasonal investigation of intertidal zones of Naozhou Island was conducted from April 2011 to January 2012. The results show that there were 64 species of macroalgae in this Sargasso field, including 15 species of Phaeophyta, 28 species of Rhodophyta, 20 species of Chlorophyta and one species of Cyanophyta, accounting for 23.44%, 43.75%, 31.25% and 1.56% of the total species, respectively. Among the 64 species, 43 species were thriving in spring, 24 species were emerging in summer, 29 species were active in autumn and 31 in winter. Only nine species were thriving throughout the whole year, including six species of Rhodophyta, two species of Chlorophyta and one species Phaeophyta, while there were 14 species living through three seasons. There were about 12-26 common species that can be found between two seasons and the species turnover rate was from 0.42 to 0.78, associated with higher turnover rates in spring and summer and lower rates in autumn and winter. Moreover, there were 13 dominant species but only Pterocladiella capillacea was the all-year dominant species, while Sargassum hemiphyllum and Corallina pilulifera were the dominant species over three seasons. Average biomass vary greatly with season, with the order of spring (848.14 g·m-2)> winter (378.57 g·m-2)> autumn (297.99 g·m-2)> summer (294.15 g·m-2). There is clearly vertical variation in the species distribution as the species number increases significantly from high tidal zone to low tidal zone, while the biomass show similar changing trend, indicating both species and biomass increase with water depth in the tidal zone. The Shannon-Weinner index was from 0.03 to 2.33, with an annual average of 1.10, while the Pielou's index was from 0.01 to 0.70, with an annual average of 036; the Margalef index varied from 0.15 to 1.65, with an annual average of 0.72; and the Simpson’s dominance index varied from 0.01 to 0.78, with an annual average of 0.39. The total water content of seaweeds changes ranged from 51.42% to 97.52%, with an average moisture of 85.21%. The total organic carbon content of seaweeds varied from 4.34% to 42.06%, with an average of 27.99%, indicating a strong carbon storage capacity. We find a clear negative correlation between the biomass of large seaweeds and dissolved inorganic nitrogen (DIN), with a coefficient of 0.49 (P<0.05). The lower DIN of seawater during spring and winter is coincided with microalgae thriving seasons.
ZHANG Caixue , ZHOU Weinan , SUN Xingli , SONG Zhiguang . Seasonal succession of macroalgae community in Naozhou Island[J]. Journal of Tropical Oceanography, 2020 , 39(1) : 74 -84 . DOI: 10.11978/2019026
表1 硇洲岛岩礁区大型海藻季节间种类更替率Tab. 1 Species subrogation ratio of macroalgae between two seasons in the rocky zone of Naozhou Island |
| 项目 | 春/夏 | 春/秋 | 春/冬 | 夏/秋 | 夏/冬 | 秋/冬 |
|---|---|---|---|---|---|---|
| 各季节种数 | 43/24 | 43/29 | 43/31 | 24/29 | 24/31 | 29/31 |
| 共有种数 | 12 | 20 | 26 | 13 | 13 | 22 |
| 种类更替率R | 0.78 | 0.62 | 0.46 | 0.68 | 0.69 | 0.42 |
表2 调查海域各季节大型海藻的优势种群Tab. 2 Dominant species of marcoalgae in each season in the study area |
| 季节 | 优势种 | 平均生物量/(g·m-2) | 生物量百分比/% | 出现频度 | 优势度 |
|---|---|---|---|---|---|
| 春季 | Sargassum hemiphyllum | 540.08 | 63.68 | 1.00 | 0.64 |
| Pterocladiella capillacea | 128.64 | 15.17 | 1.00 | 0.15 | |
| Colpomenia sinuosa | 76.33 | 9.00 | 0.80 | 0.07 | |
| Asparagopsis taxiformis | 40.94 | 4.83 | 0.60 | 0.03 | |
| 夏季 | Caulacanthus ustulatus | 49.40 | 16.79 | 0.80 | 0.13 |
| Chondracanthus intermedius | 46.08 | 15.67 | 0.40 | 0.06 | |
| Centroceras clavulatum | 35.07 | 11.92 | 0.40 | 0.05 | |
| Neogoniolithon sp. | 31.20 | 10.61 | 0.40 | 0.04 | |
| Jania sp. | 26.75 | 9.10 | 0.40 | 0.04 | |
| Cladophora stimpsonii | 20.84 | 7.09 | 1.00 | 0.07 | |
| Pterocladiella capillacea | 17.36 | 5.90 | 1.00 | 0.06 | |
| Corallina pilulifera f. sororia | 17.15 | 5.83 | 0.40 | 0.02 | |
| Caulerpa taxifolia | 16.06 | 5.46 | 0.80 | 0.04 | |
| 秋季 | Pterocladiella capillacea | 113.19 | 38.24 | 0.80 | 0.31 |
| Sargassum hemiphyllum | 55.29 | 18.68 | 0.60 | 0.11 | |
| Chondracanthus intermedius | 33.92 | 11.46 | 0.40 | 0.05 | |
| Valonia aegagropila | 29.57 | 9.99 | 0.60 | 0.06 | |
| Corallina pilulifera f. sororia | 11.45 | 3.87 | 1.00 | 0.04 | |
| 冬季 | Pterocladiella capillacea | 175.81 | 40.08 | 0.80 | 0.32 |
| Sargassum hemiphyllum | 109.51 | 24.96 | 0.60 | 0.15 | |
| Corallina pilulifera f. sororia | 47.86 | 10.91 | 1.00 | 0.11 | |
| Centroceras clavulatum | 46.60 | 10.62 | 0.60 | 0.06 |
表3 硇洲岛岩礁区大型海藻的多样性参数Tab. 3 Diversity parameters of marcoalgae in the rocky zone of Naozhou Island |
| 多样性指数 | 断 面 | 平均 | |||||
|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | R5 | |||
| 春季 | H′ | 0.03 | 1.29 | 1.45 | 1.32 | 1.19 | 1.06 |
| J | 0.02 | 0.43 | 0.34 | 0.35 | 0.37 | 0.30 | |
| d | 0.38 | 0.47 | 1.65 | 1.18 | 1.62 | 1.06 | |
| 1-λ′ | 0.01 | 0.51 | 0.58 | 0.50 | 0.31 | 0.38 | |
| 夏季 | H′ | 1.51 | 0.05 | 1.58 | 2.33 | 2.21 | 1.54 |
| J | 0.65 | 0.02 | 0.68 | 0.70 | 0.62 | 0.53 | |
| d | 0.45 | 0.64 | 0.43 | 0.76 | 0.97 | 0.65 | |
| 1-λ′ | 0.62 | 0.01 | 0.62 | 0.78 | 0.73 | 0.55 | |
| 秋季Autumn | H′ | 0.28 | 0.01 | 0.77 | 1.78 | 1.27 | 0.82 |
| J | 0.14 | 0.01 | 0.33 | 0.56 | 0.40 | 0.29 | |
| d | 0.44 | 0.15 | 0.34 | 0.73 | 0.73 | 0.48 | |
| 1-λ′ | 0.07 | 0.01 | 0.27 | 0.66 | 0.47 | 0.30 | |
| 冬季 | H′ | 0.43 | 0.75 | 0.72 | 1.95 | 1.09 | 0.99 |
| J | 0.17 | 0.37 | 0.21 | 0.61 | 0.30 | 0.33 | |
| d | 0.58 | 0.27 | 0.93 | 0.71 | 0.95 | 0.69 | |
| 1-λ′ | 0.16 | 0.29 | 0.19 | 0.67 | 0.34 | 0.33 | |
表4 硇洲岛主要大型海藻的碳含量(干重)Tab. 4 Carbon contents of major marcoalgae (dry weight) in Naozhou Island |
| 种类 | 含水率/% | TC/% | IC/% | TOC/% |
|---|---|---|---|---|
| 紫杉状海门冬Asparagopsis taxiformis | 94.94 | 35.02 | 0.00 | 35.02 |
| 哈维羽藻Bryopsis harveyana | 89.97 | 17.37 | 1.22 | 16.15 |
| 茎刺藻Caulacanthus ustulatus | 94.12 | 35.96 | 0.02 | 35.94 |
| 盾叶蕨藻Caulerpa peltata | 61.42 | 34.43 | 3.97 | 30.46 |
| 衫叶蕨藻Caulerpa taxifolia | 95.26 | 29.43 | 2.26 | 27.17 |
| 纵胞藻Centroceras clavulatum | 86.96 | 20.12 | 0.00 | 20.12 |
| 气生硬毛藻Chaetomorpha aerea | 92.11 | 36.11 | 0.27 | 35.84 |
| 中间硬毛藻Chaetomorpha media | 91.21 | 25.89 | 0.66 | 25.23 |
| 粗枝软骨藻Chondria dasyphylla | 91.09 | 26.90 | 3.63 | 23.27 |
| 簇生刚毛藻Cladophoropsis fasciculatus | 78.19 | 9.05 | 0.00 | 9.04 |
| 斯氏刚毛藻Cladophora stimpsonii | 91.55 | 22.81 | 4.66 | 18.15 |
| 囊藻Colpomenia sinuosa | 87.83 | 27.92 | 0.54 | 27.38 |
| 小珊瑚藻姐妹变型Corallina pilulifera f. sororia | 57.54 | 16.29 | 9.19 | 7.10 |
| 珊瑚藻Corallina sp. | 83.14 | 42.07 | 0.01 | 42.06 |
| 网球藻Dictyosphaeria cavernosa | 94.69 | 17.15 | 3.45 | 13.7 |
| 鹅肠菜Endarachne binghamiae | 92.13 | 36.73 | 0.01 | 36.72 |
| 条浒苔Enteromorpha clathrata | 89.17 | 26.89 | 3.09 | 23.80 |
| 肠浒苔Enteromorpha intestinalis | 93.28 | 31.25 | 1.25 | 30.00 |
| 缘管浒苔Enteromorpha linza | 97.02 | 39.12 | 0.00 | 39.12 |
| 钝乳节藻Galaxaura obtusata | 64.68 | 17.86 | 6.23 | 11.63 |
| 太平洋乳节藻Galaxaura pacifica | 74.19 | 5.46 | 0.59 | 4.86 |
| 乳节藻Galaxaura sp. | 64.68 | 17.93 | 3.41 | 14.52 |
| 小石花菜Gelidium divaricatum | 83.96 | 41.75 | 0.00 | 41.75 |
| 小衫藻Gigartina intermedia | 76.15 | 40.53 | 0.19 | 40.34 |
| 舌状蜈蚣藻Grateloupia livida | 94.46 | 35.91 | 0.00 | 35.91 |
| 繁枝蜈蚣藻Grateloupia ramosissima | 97.52 | 32.13 | 0.00 | 32.13 |
| 种类 | 含水率/% | TC/% | IC/% | TOC/% |
| 扇形拟伊藻Gymnogongrus flabelliformis | 94.09 | 37.68 | 0.00 | 37.68 |
| 羊栖菜Hizikia fusiforme | 91.55 | 37.02 | 0.27 | 36.75 |
| 冻沙菜Hypnea japonica | 86.45 | 16.61 | 1.25 | 15.36 |
| 南方团扇藻Padina australis | 89.20 | 20.18 | 0.86 | 19.32 |
| 多管藻Polysiphonia sp. | 76.82 | 20.25 | 0.00 | 20.25 |
| 拟鸡毛菜Pterocladiella capillacea | 76.54 | 41.54 | 0.73 | 40.81 |
| 亨氏马尾藻Sargassum henslowianum | 88.53 | 39.96 | 0.00 | 39.96 |
| 全缘马尾藻Sargassum integerrimum | 87.06 | 39.57 | 0.00 | 39.57 |
| 硇洲马尾藻Sargassum naozhouense | 86.48 | 38.89 | 0.00 | 38.89 |
| 丛缘叶马尾藻Sargassum duplicatum | 82.52 | 34.62 | 0.00 | 34.62 |
| 半叶马尾藻Sargassum hemiphyllum | 84.31 | 37.30 | 0.11 | 37.19 |
| 孔石莼Ulva pertusa | 95.46 | 39.50 | 0.02 | 39.48 |
| 花石莼Ulva conglobata | 85.01 | 35.28 | 1.10 | 34.18 |
| 裂片石莼Ulva fasciata | 94.80 | 39.32 | 0.00 | 39.32 |
| 石莼Ulva lactuca | 82.66 | 15.19 | 6.24 | 8.95 |
| 囊状法囊藻Valonia utricularis | 93.39 | 40.32 | 0.72 | 39.60 |
| 指状藻Valoniopsis pachynema | 51.92 | 7.66 | 3.32 | 4.34 |
| 平均值 | 85.21 | 29.37 | 1.38 | 27.99 |
表5 硇洲岛岩礁区大型海藻物种多样性与其他海域比较Tab. 5 Comparison of macroalgae diversity between reef area of Naozhou island and other seas |
| 海域 | 调查时间 | 大型海藻种类数 | 平均生物量/(g·m-2) | 多样性指数 | 均匀度 | 种类丰富度 | 数据来源 |
|---|---|---|---|---|---|---|---|
| 三亚岩相潮间带 | 2008年 | 130 | 430.86 | 1.50 | 0.64 | 0.86 | 陈自强 等(2013) |
| 大连凤鸣岛潮间带 | 2011年 | 74 | 407.07 | 1.42 | 0.88 | 0.50 | 王琳(2014) |
| 硇洲岛 | 2011年 | 64 | 454.71 | 1.10 | 0.36 | 0.72 | 本研究 |
表6 大型海藻生物量与环境因子的相关性分析Tab. 6 Correlative analysis between marcoalgae biomass and environmental factors |
| SS | WT | 盐度 | pH | DIN | $\text{PO}_{\text{4}}^{\text{3}-}$ | $\text{SiO}_{\text{3}}^{\text{2}-}$ | Chl a | TOC | 大型海藻 | |
|---|---|---|---|---|---|---|---|---|---|---|
| SS | 1 | |||||||||
| WT | -0.31 | 1 | ||||||||
| 盐度 | -0.04 | 0.20 | 1 | |||||||
| pH | 0.08 | -0.26 | -0.66** | 1 | ||||||
| DIN | -0.16 | 0.36 | -0.28 | -0.15 | 1 | |||||
| PO43- | 0.09 | -0.01 | 0.01 | -0.19 | 0.24 | 1 | ||||
| SiO32- | 0.20 | -0.47* | -0.20 | -0.04 | -0.02 | 0.10 | 1 | |||
| Chla | 0.51* | -0.23 | -0.50* | 0.38 | 0.23 | 0.09 | 0.10 | 1 | ||
| TOC | 0.88** | -0.14 | 0.16 | -0.11 | -0.11 | 0.20 | 0.14 | 0.53* | 1 | |
| 大型海藻 | 0.06 | 0.12 | 0.07 | 0.18 | -0.49* | -0.09 | -0.10 | -0.22 | 0.03 | 1 |
注: *为显著相关, p<0.05 (双尾); **为极显著相关, p<0.01 (双尾)。 |
| [1] |
蔡如星, 郑锋, 王彝豪 , 等, 1990. 舟山潮间带生态学研究——I. 种类组成及分布[J]. 东海海洋, 8(1):51-60.
|
| [2] |
陈苑 , 2011. 硇洲岛地下水水质咸化和海水入侵主要成因及对策[J]. 大众科技, (4):114-115.
|
| [3] |
陈自强, 寿鹿, 廖一波 , 等, 2013. 三亚岩相潮间带底栖海藻群落结构及其季节变化[J]. 生态学报, 33(11):3370-3382.
|
| [4] |
国家海洋环境监测中心, 2008. GB17378.4-2007 海洋监测规范第4部分: 海水分析[S]. 北京: 中国标准出版社: 57-121.
National Marine Environmental Monitoring Center, 2008. GB17378.4-2007 The specification for marine monitoring—Part 4: seawater analysis[S]. Beijing: Standards Press of China: 57-121(in Chinese).
|
| [5] |
胡兴 , 1997. 湛江硇洲岛沿岸经济海藻的种类和生态分布[J]. 怀化师专学报, 16(6):72-75.
|
| [6] |
蒋日进 , 2015. 枸杞岛近岸海域食物网结构研究[D]. 上海: 上海海洋大学: 1-118.
|
| [7] |
赖廷和, 何斌源, 黄中坚 , 等, 2019. 防城河口湾潮间带大型底栖动物群落结构研究[J]. 热带海洋学报, 38(2):67-77.
|
| [8] |
李荣冠, 江锦祥, 鲁琳 , 等, 1993. 大亚湾潮间带底栖生物种类组成与分布[J]. 海洋与湖沼, 24(5):527-535.
|
| [9] |
李夏 , 2016. 黄东海浮游植物群落结构季节和年际变化特征模拟分析[D]. 天津: 天津科技大学: 1-72.
|
| [10] |
李岩, 付秀梅 , 2015. 中国大型海藻资源生态价值分析与评估[J]. 中国渔业经济, 33(2):57-62.
|
| [11] |
罗余方 , 2017. 环境人类学视域下的海洋渔业生态环境问题研究——以粤西硇洲岛渔业生态环境变迁史为例[J]. 青海民族研究, 28(3):73-77.
|
| [12] |
马威 , 2012. 南海北部浮游植物群落的生态学研究[D]. 青岛: 中国海洋大学: 1-74.
|
| [13] |
彭欣, 谢起浪, 陈少波 , 等, 2009. 南麂列岛潮间带底栖生物时空分布及其对人类活动的响应[J]. 海洋与湖沼, 40(5):584-589.
|
| [14] |
沈国英, 黄凌风, 郭丰 , 等, 2016. 海洋生态学[M]. 3版. 北京: 科学出版社: 116-143.
|
| [15] |
施华宏 , 2001. 滨海采捕对岸带生物资源的影响及管理对策[J]. 海洋开发与管理, 18(2):32-34.
|
| [16] |
王琳 , 2014. 大连凤鸣岛潮间带大型底栖海藻的群落结构及多样性[D]. 大连: 辽宁师范大学: 1-41.
|
| [17] |
肖汉洪, 李方满 , 2003. 广东硇洲岛潮间带蟹类的组成及生态分布[J]. 贵州科学, 21(4):59-62.
|
| [18] |
杨万喜, 陈永寿 , 1996. 嵊泗列岛潮间带群落生态学研究I.岩相潮间带底栖生物群落组成及季节变化[J]. 应用生态学报, 7(3):305-309.
|
| [19] |
张华伟, 彭欣, 刘俊峰 , 等, 2018. 南麂列岛冬夏季潮间带大型底栖生物群落结构[J]. 上海海洋大学学报, 27(1):133-140.
|
| [20] |
章守宇, 孙宏超 , 2007. 海藻场生态系统及其工程学研究进展[J]. 应用生态学报, 18(7):1647-1653.
|
| [21] |
张莹 , 2016. 渤海中部浮游植物与环境因子的空间关系及季节差异分析[D]. 烟台: 中国科学院烟台海岸带研究所: 1-75.
|
| [22] |
赵素芬, 孙会强, 袁振江 , 等, 2006. 硇洲岛春季底栖海藻资源[J]. 上海水产大学学报, 15(3):321-327.
|
| [23] |
郑成兴, 李荣冠, 江锦祥 , 等, 2004. 泉州湾岩相潮间带底栖生物生态研究I. 物种多样性与分布特征[J]. 生物多样性, 12(6):594-610.
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}