
Journal of Tropical Oceanography >
Cloning and functional analysis of a new pattern recognition receptor CgLRRC69 in Pacific oyster, Crassostrea gigas
Received date: 2020-02-28
Request revised date: 2020-04-28
Online published: 2020-05-19
Supported by
National Natural Science Foundation of China(31572640)
National Natural Science Foundation of China(31572661)
Copyright
In bivalve mollusks, circulating hemocytes phagocytosis is one basic way to eliminate invading pathogens. In this study, we identified a novel Leucine-rich repeat (LRR) domain containing gene (i.e., CgLRRC69) in the Pacific oyster, Crassostrea gigas, which plays an important role in recognizing bacteria and promoting hemocyte phagocytosis. Tissue distribution analysis showed its constitutive expression in various tissues, including hemocytes, gills, muscles, mantle, heart, digestive glands, and gonads. Moreover, CgLRRC69 mRNA was significantly increased and peaked at six hours of post-infection with V. parahaemolyticus in hemocytes. Furthermore, functional assay revealed that recombinant CgLRRC69 protein can specifically bind lipopolysaccharide (LPS), indicating its function in immune defense. Meanwhile, CgLRRC69 protein can also significantly promote phagocytosis of hemocytes. Finally, CgLRRC69 depletion via RNAi in vivo resulted in obvious reduction of bacterial clearance in hemocytes. Therefore, our study demonstrates that CgLRRC69 may function as a pattern recognition receptor for Vibrio spp. and promotes hemocyte phagocytosis. This study contributes to our better understanding of the mechanism of immune defense and disease management in the Pacific oyster.
ZHANG Xiangyu , SONG Jingchen , LIU Kunna , MAO Fan , XIAO Shu , XIANG Zhiming , ZHANG Yang , YU Ziniu . Cloning and functional analysis of a new pattern recognition receptor CgLRRC69 in Pacific oyster, Crassostrea gigas[J]. Journal of Tropical Oceanography, 2021 , 40(1) : 65 -74 . DOI: 10.11978/2020024
表格1 使用的寡核苷酸引物序列Tab. 1 Sequences of oligonucleotide primers used |
| 引物 | 序列 | 用途 | |||
|---|---|---|---|---|---|
| DsGFP-F | TAATACGACTCACTATAGGNGTCAGTGGAGAGGGTGAAGG | RNAi | |||
| DsGFP-R | TAATACGACTCACTATAGGNAGGCATGGCACTCTTGAAAA | RNAi | |||
| DsLRR-F | TAATACGACTCACTATAGGNTCCGCTTGAGTTTGGATTCC | RNAi | |||
| DsLRR-R | TAATACGACTCACTATAGGNTTCCTTCAGTGAGAGGACCT | RNAi | |||
| LRRi-F | AGAACTACCCGAAGTCCTTGA | qRT-PCR | |||
| LRRi-R | CCAAACTCAAGCGGAAGACT | qRT-PCR | |||
| GAPDH-F | CTGCCAACGTGTCAGTGGTG | qRT-PCR | |||
| GAPDH-R | TCAGTGTAGCCCAGGATGCC | qRT-PCR | |||
| LRR-F | AGCATTGATAGTTGAGATCA | ORF | |||
| LRR-R | TCGAAGAGCAAATCACATGA | ORF | |||
图1 CgLRRC69完整的ORF和推导的氨基酸序列黑色字体为长牡蛎CgLRRC69的开放阅读框(ORF), 蓝色字体为翻译的氨基酸序列, 粗体表示起始密码子和终止密码子; *代表终止密码子, 负责终止肽链合成; 灰色阴影部分表示LRR-亮氨酸重复序列 Fig. 1 The complete ORF and deduced amino acid sequence of CgLRRC69. The black font is the open reading frame (ORF) of Oyster CgLRRC69, the blue font is the translated amino acid sequence, and the bold font indicates the start codon and the stop codon. * represents the stop codon, which does not encode any amino acid and is responsible for terminating the peptide chain synthesis. Shading indicates the LRR-leucine repeat sequence |
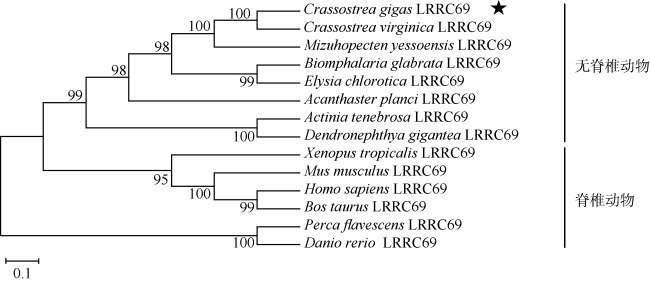
图2 长牡蛎LRRC69氨基酸序列与人(Homo sapiens)、家鼠(Mus musculus)、斑马鱼(Danio rerio)、虾夷扇贝(Mizuhopecten yessoensis)和美洲牡蛎(Crassostrea virginica)的对比图中*、:和.分别表示相同的、高度保守的和不太保守的氨基酸残基; 黑底白字表示在选取的6个物种中, 在不太保守的氨基酸残基中仍有3个以上相同氨基酸重复; 灰底黑字表示在不太保守的氨基酸残基中存在性质相似的氨基酸; GenBank登录号如下: 人(NP_001123362.1)、家鼠(XP_006538358.1)、斑马鱼(XP_005159720.1)、虾夷扇贝(XP_021346950.1)和美洲牡蛎(XP_022338017.1) Fig. 2 The amino acid sequence of Pacific oyster LRRC69, compared with Homo sapiens, Mus musculus, Danio rerio, Mizuhopecten yessoensis, and Crassostrea virginica. Identical, highly conserved and less conserved amino acid residues are indicated by *, : and . . White letters on black background indicate that in the six selected species, there are still more than three identical amino acid residues in the less conservative amino acid residues. Black letters on gray background indicate that there are amino acids with similar properties in the less conservative amino acid residues. The GenBank accession numbers are listed: Homo sapiens (NP_001123362.1), Mus musculus (XP_006538358.1), Danio rerio (XP_005159720.1), Mizuhopecten yessoensis (XP_021346950.1) and Crassostrea virginica (XP_022338017.1) |
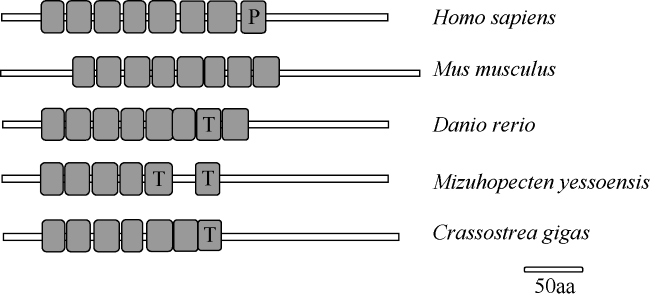
图3 LRRC69在不同物种中的结构图解P表示Pfam: LRR, T表示LRR-TYP, 空白表示普通LRR; GenBank登录号如下: 人(NP_001123362.1)、家鼠(XP_006538358.1)、斑马鱼(XP_005159720.1)、虾夷扇贝(XP_021346950.1)、美洲牡蛎(XP_022338017.1) Fig. 3 Schematic of LRRC69 structures in different species. P means Pfam: LRR; T means LRR-TYP; blank means ordinary LRR. The GenBank accession numbers are listed: Homo sapiens (NP_001123362.1), Mus musculus (XP_006538358.1), Danio rerio (XP_005159720.1), Mizuhopecten yessoensis (XP_021346950.1) and Crassostrea virginica (XP_022338017.1) |
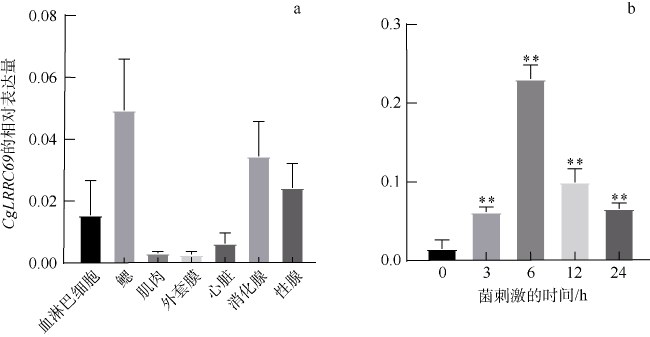
图5 CgLRRC69基因的相对表达量a. 长牡蛎组织中CgLRRC69的分布, 包括血淋巴细胞、鳃、外套膜、肌肉、心脏、消化腺和性腺; b. CgLRRC69在细菌刺激下的表达变化, 数值以Mean ± SEM表示(N=3), **表示p<0.01, 即差异极显著 Fig. 5 CgLRRC69 gene expression in tissues and under bacterial challenge. (a) Tissue distribution of CgLRRC69 in oyster, including hemocytes, gill, muscle, mantle, heart, digestive gland, and gonads. (b) Variation in expression of CgLRRC69 in response to bacterial challenge. Data are expressed as Mean ± SEM (N=3). ** means p<0.01, namely, the difference is extremely significant |
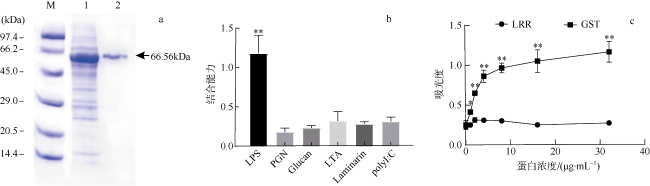
图6 CgLRRC69蛋白的体外表达及对LPS的亲和力特异性分析a. 经SDS-PAGE分析纯化后的CgLRRC69蛋白, 用考马斯亮蓝染色显示; M为分子量标记; 数字1表示泳道1, 为总细菌蛋白; 数字2表示泳道2, 为纯化的蛋白质。b. 测定病原菌相关的模式识别分子与CgLRRC69蛋白的结合亲力, **表示p<0.01, 即差异极显著。c. 酶联免疫吸附法测定LRR结构域蛋白与固定化LPS的结合, 数值以Mean ± SEM表示(N=3); *表示p<0.05, 即差异显著; **表示p<0.01, 即差异极显著 Fig. 6 CgLRRC69 protein with affinity specificity for LPS. (a) The purified CgLRRC69 recombinant protein was analyzed by SDS-PAGE and visualized with Coomassie brilliant blue staining. M: Molecular weight marker; Lane 1: total bacterial protein; Lane 2: purified protein. (b) PAMPs binding affinity with CgLRRC69. ** means p<0.01, namely, the difference is extremely significant. (c) The binding of LRR domain protein to immobilized LPS as determined by Enzyme-linked immunosorbent assay. Data are expressed as Mean ± SEM (N=3). * means p<0.05, namely, the difference is significant; ** means p<0.01, namely, the difference is extremely significant |
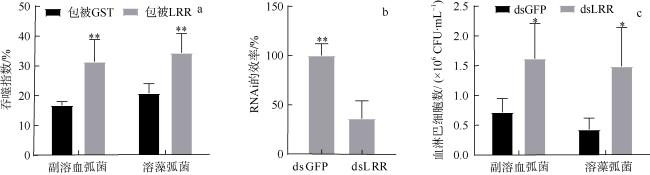
图7 CgLRRC69在吞噬和杀菌中的作用a. CgLRRC69重组蛋白对血淋巴吞噬细菌的影响, **表示p<0.01, 即差异极显著; b. RNAi敲降效率, **表示p<0.05, 即差异极显著; c. 对照组与dsCgLRRC69敲降处理组杀菌能力的差异, 数值以Mean ± SEM表示(N=3); *表示p<0.01, 即差异显著 Fig. 7 The role of CgLRRC69 in phagocytosis and sterilization. (a) The effects of CgLRRC6 recombinant protein coating on the phagocytic ability of hemocytes. ** means p<0.01, namely, the difference is extremely significant. (b) dsCgLRRC69 knockdown efficiency. Experimental group: dsLRRC69; control group: dsGFP. ** means p<0.01, namely, the difference is extremely significant. (c) Display of the difference in bactericidal ability between the control group and dsCgLRRC69 knockdown treatment group. Data are expressed as Mean ± SEM (N=3). * means p<0.05, namely, the difference is significant |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}