
Journal of Tropical Oceanography >
Cloning and expression analysis of aquaporin gene AQP4 cDNA from Pinctada fucata martensii
Received date: 2019-08-19
Request revised date: 2019-11-28
Online published: 2020-05-19
Supported by
The Earmarked Fund for Modern Agro-industry Technology Research System(CARS-49)
Marine Ecological Civilization Project(ISEE2018PY03)
Science and Technology Planning Project of Guangdong Province, China(2017B0303014052)
Copyright
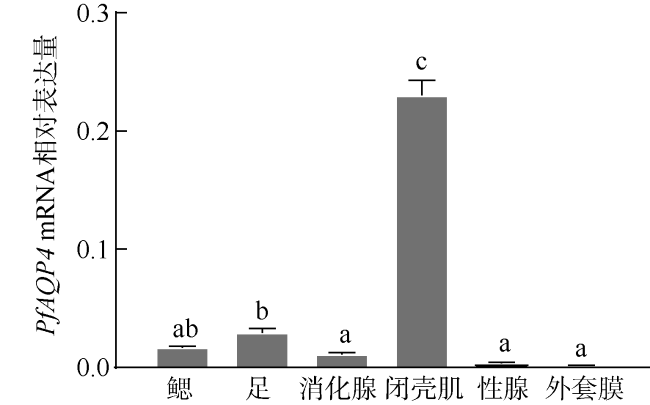
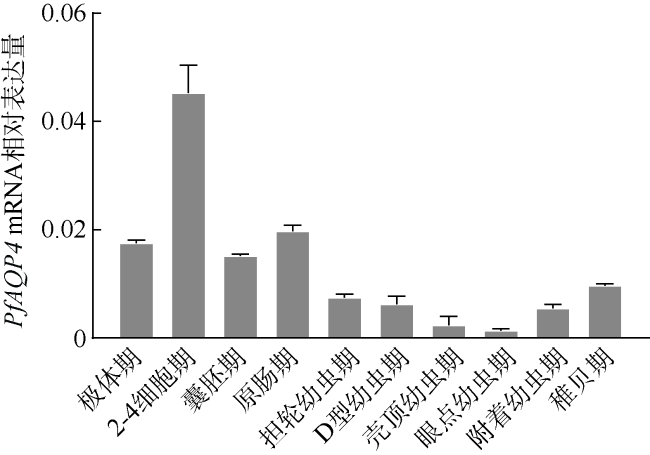
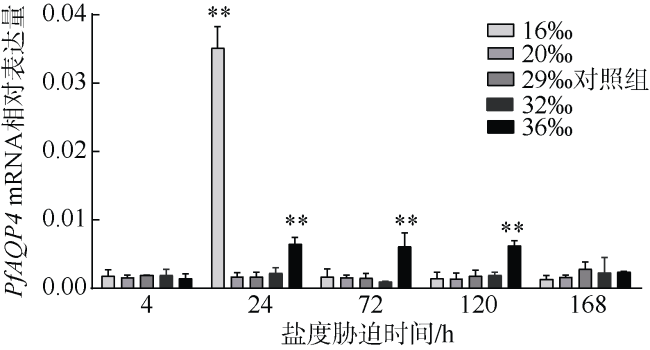
Aquaporin 4 (AQP4) is a major channel that passively transports water. In this study, based on a fragment of AQP4 of Pinctada fucata martensii obtained by transcriptome sequencing, the full cDNA was obtained using RACE technology, named PfAQP4, which includes 114 bp of 5'UTR, 839 bp of 3'UTR, and 858 bp of the Open Reading Frame (ORF) encoding a total of 285 amino acids. The PfAQP4 has six alpha-helix transmembrane regions, five loops, one major intrinsic protein (MIP) domain, and two NPAs (Asn-Pro-Ala) motif, indicating that it belongs to AQP1-like type. Real-time fluorescent quantitative PCR was used to analyze the mRNA expression pattern of PfAQP4 in different tissues, different developmental stages, and different salinity stress conditions. The results showed that: (1) PfAQP4 was expressed in all tested tissues, and its expression level was higher in adductor muscle, foot, and gill; (2) in different developmental stages, PfAQP4 was increased first and then decreased, highly expressed in the 2-4 cell stage and lowly expressed in eye-spot larva stage; (3) the expression of gill PfAQP4 mRNA in hypersaline group (36‰) increased significantly at 24h, 72h, and 120h, and returned to the control level at 168 h; in the hyposaline group (16‰), PfAQP4 was significantly up-regulated at 24 h, and the expression level returned to the control level at 72h. These results showed that the salinity can affect the expression of PfAQP4 in gill, and PfAQP4 had a very important effect on the osmoregulation of P. f. martensii.
Key words: Pinctada fucata martensii; aquaporin 4; salinity stress; osmoregulation
PAN Xiaolan , LIU Huiru , XU Meng , XU Hanzhi , ZHANG Hua , HE Maoxian . Cloning and expression analysis of aquaporin gene AQP4 cDNA from Pinctada fucata martensii[J]. Journal of Tropical Oceanography, 2020 , 39(3) : 66 -75 . DOI: 10.11978/2019074
表1 本研究所用的引物序列Tab. 1 Primers used in this study |
| 引物名称 | 引物序列(5′—3′) | 注释 |
|---|---|---|
| GSP5 | CGATAGCCCAAAGGCAAAAGATA | 5′RACE外引物 |
| NGSP5 | GAAGGTGGCAGAGGTACTCTTGAAGG | 5′RACE内引物 |
| GSP3 | GTTCGTCTTTGCTGCTCCTTCC | 3′RACE外引物 |
| NGSP3 | GACCTGAAGAAGGACGAAAACG | 3′RACE内引物 |
| Faqp | CTGTGATGGCGTCAACTGATG | PfAQP4定量引物 |
| Raqp | CGCAATCGTGAGAAGATGACC | |
| F18S | CGTTTCAACAAGACGCCAGTAG | 内参基因18S定量引物 |
| R18S | ACGAAAAAAAGGTTTGAGAGACG |
图1 PfAQP4 的 cDNA全长序列及推导的氨基酸小写字母表示5′UTR和3′UTR序列, 大写字母表示ORF序列, 阴影部分表示SMART预测的MIP结构域, 方框示NPA序列, 下划线示6个跨膜区, *代表蛋白翻译终止子, 粗体aataaa代表polyA加尾信号 Fig. 1 cDNA sequence and deduced amino acid sequence of PfAQP4. Lower case letters indicate 5'UTR and 3'UTR, upper case letters indicate ORF, shaded areas indicate SMART-predicted MIP domains, boxes indicate NPA sequences, underlined letters indicate six transmembrane regions, and * represents protein translation terminator. aataaa in bold represents polyadenylation tail signal |
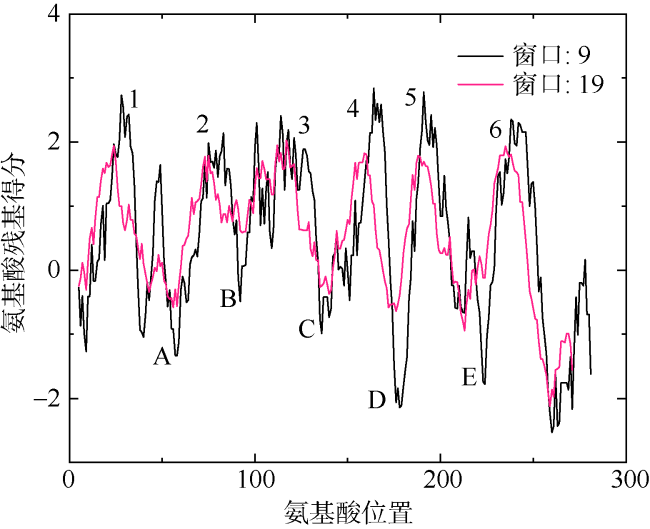
图2 PfAQP4疏水性示意图1—6表示6个跨膜区; A—E表示5个环; 窗口大小是指用于轮廓计算的间隔长度, 在计算给定残基i的得分时, 考虑以残基i为中心的选定长度区间内的氨基酸 Fig. 2 Hydrophobic plots of PfAQP4. Numbers 1-6 represents six transmembrane regions, A-E represent five loops, and the windows size refers to the interval length used for contour calculation. When calculating the score of a given residue i, the amino acids in the selected length interval centered on residue i are considered |
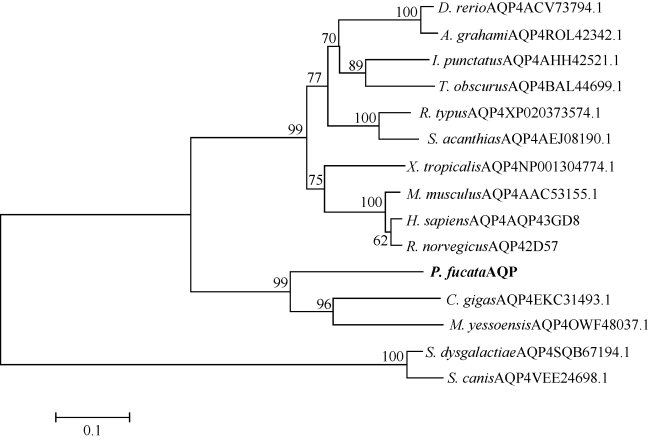
表2 PfAQP4与其他已知AQP4同源序列氨基酸一致性比较Tab. 2 Amino acid identity comparison of PfAQP4 with other known AQP4 homologues |
| 物种名 | 多序列比对用缩写名 | 登录号 | 一致性/% |
|---|---|---|---|
| 马氏珠母贝(Pinctada fucata martensii) | P.f | MN310685 | 100 |
| 斑马鱼(Danio rerio) | D.r | ACV73794.1 | 39.5 |
| 鱇浪白鱼(Anabarilius grahami) | A.g | ROL42342.1 | 37.9 |
| 斑点叉尾鮰(Ictalurus punctatus) | I.p | AHH42521.1 | 40.9 |
| 暗纹东方鲀(Takifugu obscurus) | T.o | BAL44699.1 | 40.3 |
| 小鼠(Mus musculus) | M.m | AAC53155.1 | 40.4 |
| 大鼠(Rattus norvegicus) | R.n | 2D57 | 41.1 |
| 智人(Homo sapiens) | H.s | 3GD8 | 42.2 |
| 非洲爪蟾(Xenopus tropicalis) | X.t | NP_001304774.1 | 39.9 |
| 长牡蛎(Crassostrea gigas) | C.g | EKC31493.1 | 53.4 |
| 虾夷扇贝(Mizuhopecten yessoensis) | M.y | OWF48037.1 | 52.2 |
| 停乳链球菌(Streptococcus dysgalactiae) | SQB67194.1 | 23.4 | |
| 犬链球菌(Streptococcus canis) | VEE24698.1 | 23.5 |
图4 PfAQP4与其他物种AQP4的多序列比对α1—α6为6个跨膜区, 方框表示NPA基序, 黑色背景白字表示纵列碱基保守性≥50%, 灰色表示纵列各碱基相似, 白色背景黑字表示碱基不保守 Fig. 4 Multi-sequence alignment of PfAQP4 with other species AQP4. α1-α6 represents six transmembrane regions, the box represents the NPA motif, the white letter with black background indicates the column are conserved (≥50%), the gray shading indicates the columns are similar, and the black letter with white background indicates that the residues are not conserved |
| 1 |
安皓 , 2017. 水通道蛋白7、8及精液pH值对大菱鲆(Scophthalmus maximus)精子活力影响研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所).
|
| 2 |
陈海 , 2005. AQP4、AQP9在大鼠眼组织的定位与功能研究[D]. 重庆: 重庆医科大学.
|
| 3 |
丁颖 , 2014. 高渗钠对体外培养星形胶质瘤U251细胞膜AQP4内化的影响研究[D]. 长沙: 中南大学.
|
| 4 |
高沿 , 2016. 凡纳滨对虾水孔蛋白-4基因克隆、表达及其对盐度胁迫的反应研究[D]. 北京: 中国科学院大学.
|
| 5 |
何毛贤, 姜卫国, 潘金培 , 2002. CB抑制合浦珠母贝受精卵第一极体释放的染色体分离[J]. 水产学报, 26(1):15-20.
|
| 6 |
林浩然 , 1999. 鱼类生理学[M]. 广州: 广东高等教育出版社: 193-226.
|
| 7 |
沈亦平, 姜海波, 刘汀 , 等. 1993. 合浦珠母贝卵子成熟的细胞学观察[J]. 武汉大学学报(自然科学版), ( 5):109-114.
|
| 8 |
水柏年 , 2007. 若干环境因子对缢蛏Sinonovacula constricta(Lamarck)快速净化的影响研究[J]. 现代渔业信息, 22(9):6-10.
|
| 9 |
万茜 , 2014. 香港牡蛎( Crassostrea hongkongensis)水通道蛋白基因家族的克隆与表达分析[D]. 北京: 中国科学院大学.
|
| 10 |
王吉桥, 靳翠丽, 姜静颖 , 2001. 不同降盐度方式对海湾扇贝幼贝存活的影响[J]. 中国水产科学, 8(3):35-39.
|
| 11 |
王如才, 王昭萍, 张建中 , 1993. 海水贝类养殖学[M]. 青岛: 青岛海洋大学出版社: 213.
|
| 12 |
王劭雯 , 2012. 皱纹盘鲍幼鲍对海水盐度的耐受性分析[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
| 13 |
张鑫磊, 陈四清, 刘寿堂 , 等, 2006. 温度、盐度对半滑舌鳎胚胎发育的影响[J]. 海洋科学进展, 24(3):342-348.
|
| 14 |
赵明, 陈超, 柳学周 , 等, 2011. 盐度对七带石斑鱼胚胎发育和卵黄囊仔鱼生长的影响[J]. 渔业科学进展, 32(2):16-21.
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}