
Journal of Tropical Oceanography >
Progress on the community distribution and ecological functions of nitrite-oxidizing bacteria
Copy editor: YIN Bo
Received date: 2020-05-06
Request revised date: 2020-05-22
Online published: 2020-06-01
Supported by
National Natural Science Foundation of China(9185111)
National Natural Science Foundation of China(31870100)
Natural Science Foundation of Guangdong Province(2019B1515120066)
Copyright
Microbe-mediated nitrification is an essential part of global nitrogen cycle; and nitrite-oxidizing bacteria (NOB), which catalyze the second step of nitrification, have received more and more attention recently. Up to date, those isolated NOB have been identified as four phyla, belonging to seven genera. Among them, Nitrospira bacteria become a hotspot for research due to the high diversity and wide ecological distribution. In this paper, we reviewed the composition, phylogenetic relationship, nitrite oxidizing and carbon fixation pathways of NOB. We also emphasized the functional importance on global nitrogen and carbon of NOB, and highlighted research prospect for NOB.
HONG Yiguo , JIAO Lijing , WU Jiapeng , LONG Aimin , WANG Wei . Progress on the community distribution and ecological functions of nitrite-oxidizing bacteria[J]. Journal of Tropical Oceanography, 2021 , 40(2) : 139 -146 . DOI: 10.11978/2020043
图1 不同种类亚硝酸盐氧化细菌的系统进化关系[根据Daims等(2016)绘制]图中括号内的数字表示某个门或者属的亚硝酸盐氧化细菌的包含的种的个数; 内环表示亚硝酸盐氧化细菌在门分类水平上的种类和相对丰度; 外环表示亚硝酸盐氧化细菌在属分类水平上的种类和相对丰度 Fig. 1 Phylogenetic tree among different species of nitrite oxidizing bacteria |
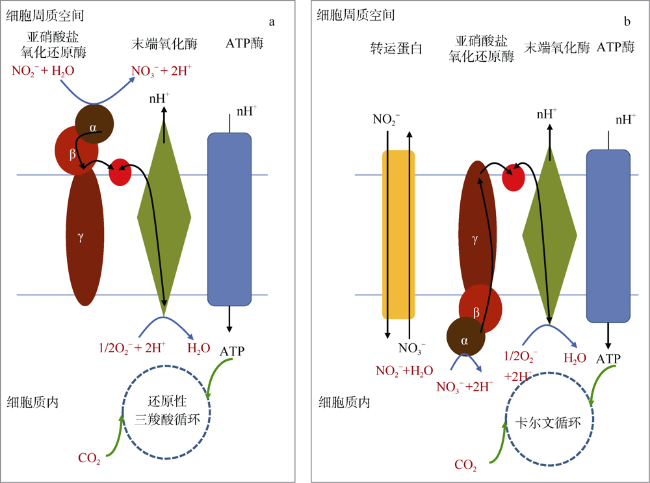
图2 亚硝酸盐氧化细菌两种不同的亚硝酸盐氧化和自养固碳模型a. 第一种模型: 在周质空间氧化亚硝酸盐偶联还原性三羧酸循环自养固碳; b. 第二种模型: 在细胞质内氧化亚硝酸盐偶联卡尔文循环自养固碳 Fig. 2 Two different modes of nitrous oxide coupled to autotrophic carbon sequestration by nitrous oxide bacteria (NOB) The first mode is to oxidize nitrite coupled with reductive tricarboxylic acid in the periplasmic space, and the second one is to oxidize nitrite coupled with Calvin cycle in the cytoplasm |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}