
Journal of Tropical Oceanography >
Study on stable isotopes of carbon and nitrogen of main fishery organisms in the southwestern waters of Daya Bay, South China Sea in winter 2020
Copy editor: LIN Qiang
Received date: 2021-08-25
Revised date: 2021-12-02
Online published: 2021-12-13
Supported by
National Key Research and Development Program of China(2018YFC1407501)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0402)
Guangdong Provincial Science and technology plan project(2020B1212060001)
Central Public-interest Scientific Institution Basal Research Fund, CAFS(2020TD15)
Central Public-interest Scientific Institution Basal Research Fund, South China Sea Fisheries Research Institute, CAFS(2021SD03)
Financial Fund of the Ministry of Agriculture and Rural Affairs, P.R. China(NFZX2021)
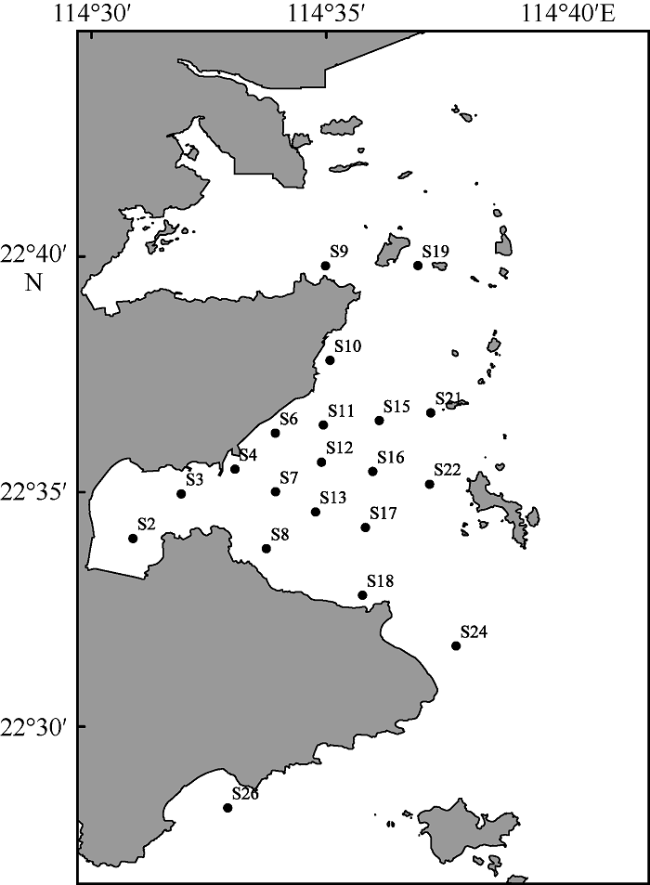
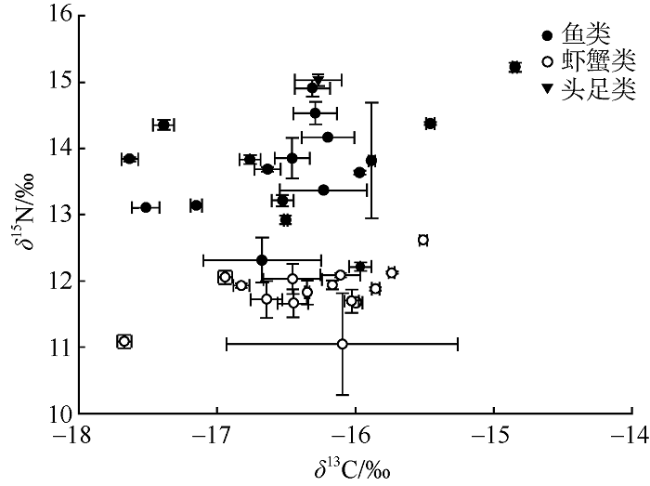
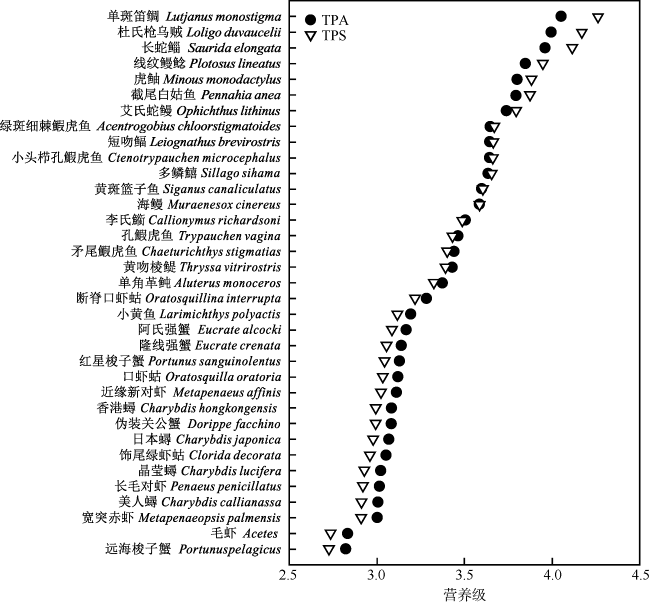
To understand the trophic structure of food webs in the southwestern waters of Daya Bay, 35 fishery organisms were collected by bottom trawl in January 2020, and the stable isotopes of carbon and nitrogen (δ13C and δ15N) of the fishery organisms were analyzed. Based on the stable isotopes of carbon and nitrogen, six quantitative ecological indicators for nutritional structure of the food web and the trophic levels of the main fishery organisms were calculated; and a continuous nutrition spectrum was made. Most of the fishery organisms captured were fish, crabs and shrimp. The δ13C and δ15N values were -17.63‰ to -14.85‰ and 12.92‰ to 15.46‰ for fishes, respectively, with average values being -16.47‰ and 13.80‰. They were -17.67‰ to -15.51‰ and 11.05‰ to 12.62‰ for shrimp and crabs, with average values of -16.30‰ and 11.85‰, respectively. According to the value of δ15N, the trophic levels of the main fishery organisms were calculated using the additive model (trophic position by the additive model, TPA) and the scaled model (trophic position by the scaled model, TPS). The result showed that there was no significant differences between the results calculated by the two models (P > 0.1), and the average trophic level of fish was higher than that of shrimp and crabs. This study indicates the food web in the southwestern waters of Daya Bay is characterized by a relatively simple initial food source, few trophic levels, short food chain, low trophic level diversity, and high degree of redundancy in trophic structure. The abundance of organisms with high trophic levels decreased, and the structure of the food web shifted from complex to simplistic, and the ecosystem became less stable in the past 30 years. The study adds new basic information about the food web structure in the study area. It also provides a reference for protecting the fishery resources of Daya Bay and maintaining the stability of the ecosystem structure there.
ZHANG Wanru , LIU Qingxia , HUANG Honghui , QIN Xiaoqing , LI Jiajun , CHEN Jianhua . Study on stable isotopes of carbon and nitrogen of main fishery organisms in the southwestern waters of Daya Bay, South China Sea in winter 2020[J]. Journal of Tropical Oceanography, 2022 , 41(3) : 147 -155 . DOI: 10.11978/2021108
表1 大亚湾西南海域主要渔业生物的碳稳定同位素比值、氮同位素比值、长度范围和营养级(n=2)Tab. 1 Stable isotope ratios of δ13C and δ15N, length range and trophic level (TL) of major fishery organisms in the southwestern waters of Daya Bay (n=2) |
| 种类 | δ13C/‰ | δ15N/‰ | 长度范围/mm | 营养级 | ||
|---|---|---|---|---|---|---|
| TPA | TPS | |||||
| 鱼类 | 多鳞鱚 Sillago sihama | -15.88 | 13.82 | 120~170 | 3.64 | 3.66 |
| 单角革鲀 Aluterus monoceros | -16.50 | 12.92 | 3.37 | 3.32 | ||
| 黄斑篮子鱼 Siganus canaliculatus | -16.63 | 13.69 | 167~176 | 3.60 | 3.61 | |
| 小头栉孔鰕虎鱼 Ctenotrypauchen microcephalus | -16.76 | 13.83 | 65~69 | 3.64 | 3.66 | |
| 小黄鱼 Larimichthys polyactis | -16.67 | 12.31 | 116~148 | 3.19 | 3.12 | |
| 绿斑细棘鰕虎鱼 Acentrogobius chloorstigmatoides | -16.46 | 13.85 | 3.65 | 3.67 | ||
| 孔鰕虎鱼 Trypauchen vagina | -16.53 | 13.22 | 89~100 | 3.46 | 3.43 | |
| 矛尾鰕虎鱼 Chaeturichthys stigmatias | -17.15 | 13.14 | 94~178 | 3.44 | 3.40 | |
| 虎鲉 Minous monodactylus | -15.46 | 14.38 | 3.80 | 3.88 | ||
| 黄吻棱鳀 Thryssa vitrirostris | -17.52 | 13.11 | 3.43 | 3.39 | ||
| 李氏䲗 Callionymus richardsoni | -16.23 | 13.37 | 70~91 | 3.51 | 3.49 | |
| 海鳗 Muraenesox cinereus | -15.97 | 13.64 | 98~120 | 3.58 | 3.59 | |
| 短吻鲾 Leiognathus brevirostris | -17.63 | 13.84 | 44~59 | 3.64 | 3.67 | |
| 截尾白姑鱼 Pennahia anea | -17.39 | 14.36 | 145 | 3.80 | 3.88 | |
| 线纹鳗鲶 Plotosus lineatus | -16.29 | 14.53 | 21~215 | 3.85 | 3.95 | |
| 长蛇鲻 Saurida elongata | -16.31 | 14.91 | 185~233 | 3.96 | 4.12 | |
| 艾氏蛇鳗 Ophichthus lithinus | -16.20 | 14.17 | 185~260 | 3.74 | 3.80 | |
| 单斑笛鲷 Lutjanus monostigma | -14.85 | 15.23 | 4.05 | 4.26 | ||
| 虾蟹类 | 远海梭子蟹 Portunus pelagicus | -16.09 | 11.05 | 2.82 | 2.73 | |
| 红星梭子蟹 Portunus sanguinolentus | -16.11 | 12.09 | 3.13 | 3.04 | ||
| 晶莹蟳 Charybdis lucifera | -16.64 | 11.72 | 3.02 | 2.93 | ||
| 伪装关公蟹 Dorippe facchino | -16.83 | 11.93 | 3.08 | 2.99 | ||
| 香港蟳 Charybdis hongkongensis | -16.17 | 11.94 | 3.08 | 3.00 | ||
| 隆线强蟹 Eucrate crenata | -15.73 | 12.12 | 3.14 | 3.05 | ||
| 美人蟳 Charybdis callianassa | -15.99 | 11.67 | 3.01 | 2.91 | ||
| 阿氏强蟹 Eucrate alcocki | -15.96 | 12.22 | 3.17 | 3.09 | ||
| 日本蟳 Charybdis japonica | -15.85 | 11.88 | 3.07 | 2.98 | ||
| 口虾蛄 Oratosquilla oratori | -16.94 | 12.06 | 3.12 | 3.04 | ||
| 饰尾绿虾蛄 Clorida decorata | -16.35 | 11.83 | 3.05 | 2.96 | ||
| 断脊口虾蛄 Oratosquillina interrupta | -15.51 | 12.62 | 3.28 | 3.22 | ||
| 近缘新对虾 Metapenaeus affinis | -16.45 | 12.03 | 3.11 | 3.03 | ||
| 长毛对虾 Penaeus penicillatus | -16.03 | 11.70 | 3.01 | 2.92 | ||
| 宽突赤虾 Metapenaeopsis palmensis | -16.45 | 11.66 | 3.00 | 2.91 | ||
| 毛虾 Acetes | -17.67 | 11.08 | 2.83 | 2.73 | ||
| 头足类 | 杜氏枪乌贼 Loligo duvaucelii | -16.27 | 15.03 | 30~200 | 3.99 | 4.17 |
表2 营养结构分析度量参数Tab. 2 Measurement parameters of trophic structure analysis |
| 参数名称 | 简写 | 含义 | 参数值 |
|---|---|---|---|
| δ13C比值范围 | CR | 图中δ13C 最大值与最小值之间的绝对距离, 反映初始食源的多样性特征 | 2.83 |
| δ15N比值范围 | NR | 图中δ15N 最大值与最小值之间的绝对距离, 反映消费者所占的营养层次 | 4.18 |
| 多边形面积 | TA | 由所有物种在δ13C-δ15N 双位图上组成的多边形面积, 是衡量群落占据的生态位空间的总数量, 代表食物网中营养多样性的总范围 | 8.30 |
| 质心均距 | CD | 质心坐标为所有样品δ13C、δ15N的平均值, 计算所有样品到质心的欧氏距离, 并得出平均值, 表示群落营养多样性的平均水平 | 1.18 |
| 最短均值 | NND | 相邻两点间最短距离的平均值, 反映群落的聚集度密度 | 0.31 |
| 最短均值标准差 | SDNND | 相邻两点间最短距离的标准差, 反映群落物种聚集度均匀度, 表示营养生态位分布范围水平 | 0.23 |
表3 典型海域食物网营养结构Tab. 3 Trophic structure of food web in typical sea areas |
| 区域 | 参考文献 | CR | NR | TA | CD | NND | SDNND |
|---|---|---|---|---|---|---|---|
| 大亚湾西南海域 | 本文 | 2.83 | 4.18 | 8.30 | 1.18 | 0.31 | 0.23 |
| 海陵湾 | 张文博 等, 2019 | 2.44 | 3.06 | 5.10 | 0.93 | 0.55 | 0.35 |
| 陵水湾 | 张文博 等, 2019 | 4.45 | 4.66 | 11.18 | 1.49 | 0.60 | 0.54 |
| 珠江口海域 | 曾艳艺 等, 2018 | 12.58 | 7.57 | — | — | — | — |
| 海州湾 | 谢斌 等, 2017 | 3.70 | 6.40 | 13.00 | 1.37 | 0.61 | 0.64 |
| 南海中西部海域 | 黄佳兴 等, 2019 | 3.49 | 4.91 | 9.48 | 1.20 | 1.69 | 0.74 |
| 南沙群岛西南陆架区 | 宁加佳 等, 2016 | 3.40 | 4.30 | 5.80 | 1.00 | 0.37 | 0.43 |
| 东海中北部海域 | 纪炜炜 等, 2015 | 3.38 | 3.45 | 7.02 | 1.12 | 0.42 | 0.28 |
| [1] |
贡艺, 陈玲, 李云凯, 2017. 海洋生态系统稳定同位素基线的选取[J]. 应用生态学报, 28(7): 2399-2404.
|
| [2] |
黄佳兴, 龚玉艳, 徐姗楠, 等, 2019. 南海中西部渔场主要渔业生物碳氮稳定同位素特征[J]. 热带海洋学报, 38(1): 76-84.
|
| [3] |
黄小平, 黄良民, 宋金明, 等, 2019. 营养物质对海湾生态环境影响的过程与机理[M]. 北京: 科学出版社: 549-558. (in Chinese)
|
| [4] |
纪炜炜, 李圣法, 陈雪忠, 等, 2015. 基于稳定同位素方法的东海北部及其邻近水域主要游泳动物营养结构变化[J]. 海洋渔业, 37(6): 494-500.
|
| [5] |
牟新悦, 陈敏, 张琨, 等, 2017. 夏季大亚湾悬浮颗粒有机物碳、氮同位素组成及其物源指示[J]. 海洋学报, 39(2): 39-52.
|
| [6] |
宁加佳, 杜飞雁, 王雪辉, 等, 2016. 南沙群岛西南部陆架区底层鱼类营养结构研究[J]. 海洋与湖沼, 47(2): 468-475.
|
| [7] |
王友绍, 王肇鼎, 黄良民, 2004. 近20年来大亚湾生态环境的变化及其发展趋势[J]. 热带海洋学报, 23(5): 85-95.
|
| [8] |
王玉堃, 2015. 耳石微细结构和微化学示踪技术在鱼类种群生态学研究中的应用[D]. 青岛: 中国海洋大学.
|
| [9] |
谢斌, 李云凯, 张虎, 等, 2017. 基于稳定同位素技术的海州湾海洋牧场食物网基础及营养结构的季节性变化[J]. 应用生态学报, 28(7): 2292-2298.
|
| [10] |
徐军, 张敏, 谢平, 2010. 氮稳定同位素基准的可变性及对营养级评价的影响[J]. 湖泊科学, 22(1): 8-20.
|
| [11] |
徐姗楠, 郭建忠, 范江涛, 等, 2020. 夏季大亚湾鱼类群落结构与多样性[J]. 生态学杂志, 39(4): 1254-1264.
|
| [12] |
杨文超, 黄道建, 陈继鑫, 等, 2020. 大亚湾近十年沉积物中汞、砷分布及污染评价[J]. 水产科学, 39(6): 915-921.
|
| [13] |
曾艳艺, 赖子尼, 杨婉玲, 等, 2018. 珠江河口渔业生物稳定同位素营养级分析[J]. 生态学杂志, 37(1): 194-202.
|
| [14] |
张硕, 高世科, 于雯雯, 等, 2019. 碳、氮稳定同位素在构建海洋食物网及生态系统群落结构中的研究进展[J]. 水产养殖, 40(7): 6-10.
|
| [15] |
张文博, 黄洪辉, 李纯厚, 等, 2019. 华南典型海湾主要渔业生物碳氮稳定同位素研究[J]. 南方水产科学, 15(5): 9-14.
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}