
Journal of Tropical Oceanography >
Oyster hydrolysates alleviate 5-fluorouracil-induced intestinal mucosal injury on S180 tumor-bearing mice*
Received date: 2021-11-10
Revised date: 2021-12-21
Online published: 2021-12-22
Supported by
National Key Research and Development Program of China(2018YFC0311202)
Marine Economy Development Special Project of Guangdong Province(GDNRC[2020]038)
Marine Economy Development Special Project of Guangdong Province(GDNRC[2020]036)
National Natural Science Foundation of Guangdong, China(2018A030313088)
Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou)(GML2019ZD0406)
Key-Area Research and Development Program of Guangdong Province(2020B1111030004)
Innovation Academy of South China Sea Ecology and Environmental Engineering, Chinese Academy of Sciences(ISEE2021PY05)
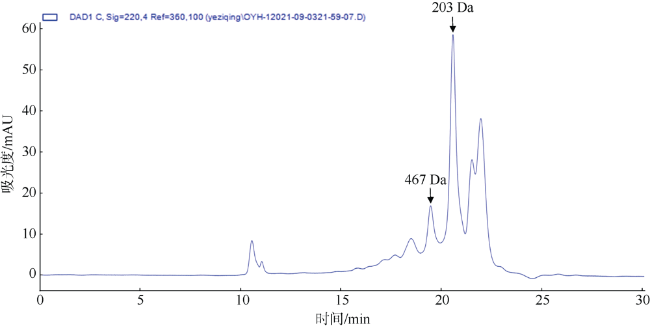
In this study, oyster meat was used as the raw material to produce trypsin hydrolysates of oyster (OYH). The amino acid composition and relative molecular weight distribution of OYH were then analyzed. The effect of OYH on reducing 5-fluorouracil (5-FU) induced intestinal mucosal injury on S180 tumor-bearing mice was preliminarily investigated. The results showed that the essential amino acids in the total amino acids of OYH accounted for 36.44 %, and content of glutamic acid was the highest. The molecular weight of OYH was mostly below 1 kDa, accounting for 92.18 %. The results showed that the tumor inhibition ratio of 5-FU group was 31.00 %. Compared with model group, the tumor inhibition ratio of mice in OYH-L+5-FU, OYH-M+5-FU and OYH-H+5-FU group were significantly increased (p < 0.001) with the percentage of 38.24 %, 46.27 % and 47.57 %, respectively. Tumor inhibition ratios of OYH-M+5-FU and OYH-H+5-FU group were greatly higher than 5-FU (p < 0.01, p < 0.001). Meanwhile, OYH can alleviate intestinal mucosal injury caused by 5-FU chemotherapy. Compared with 5-FU alone treatment, OYH-L+5-FU, OYH-M+5-FU and OYH-H+5-FU group significantly increased the villus height, crypt depth, mucosa thickness and villus surface area of small intestine (p < 0.01, p < 0.001), which indicated that OYH can improve the state of intestinal crypt and epithelial villi, maintain the integrity of the intestinal mucosal barrier during chemotherapy. In addition, compared with model group, the percentage of neutrophils was decreased but the percentage of lymphocytes was increased in OYH-L+5-FU, OYH-M+5-FU and OYH-H+5-FU group (p < 0.05, p < 0.01). And the OYH-H+5-FU group can largely increase the levels of total protein, albumin and immune globulin in serum (p < 0.05, p < 0.01, p < 0.001), which showed that OYH play a role in the immunologic function, nutritional improvement and level of immune antibody regulation in serum immunoregulation. In conclusion, trypsin hydrolysates of oyster meat have the potential for becoming an adjuvant nutritional food in the process of chemotherapy, protecting intestinal mucosa and improving body tolerance.
YE Ziqing , ZHAO Xiangdan , CAI Bingna , WAN Peng , CHEN Hua , PAN Jianyu . Oyster hydrolysates alleviate 5-fluorouracil-induced intestinal mucosal injury on S180 tumor-bearing mice*[J]. Journal of Tropical Oceanography, 2022 , 41(5) : 105 -116 . DOI: 10.11978/2021155
表1 动物不同试验分组及处理方式Tab. 1 Different experimental groups and treatments of animals |
| 组名 | 是否移植肉瘤细胞株 | 腹腔注射5-氟尿嘧啶剂量/(mg·kg-1) | 每日灌胃牡蛎酶解物剂量/(mg·kg-1) |
|---|---|---|---|
| 正常组 | 否 | — | — |
| 模型组 | 是 | — | — |
| 5-氟尿嘧啶组(5-FU组) | 是 | 30 | — |
| 低剂量牡蛎胰蛋白酶水解物联合5-氟尿嘧啶组(OYH-L+5-FU组) | 是 | 30 | 200 |
| 中剂量牡蛎胰蛋白酶水解物联合5-氟尿嘧啶组(OYH-M+5-FU组) | 是 | 30 | 400 |
| 高剂量牡蛎胰蛋白酶水解物联合5-氟尿嘧啶组(OYH-H+5-FU组) | 是 | 30 | 800 |
注: —表示未做处理 |
表3 OYH的氨基酸组成分析Tab. 3 Analysis of amino acid composition of OYH |
| 氨基酸种类 | 氨基酸含量/% | 氨基酸种类 | 氨基酸含量/% |
|---|---|---|---|
| 天冬氨酸 Asp | 3.76 | 亮氨酸 Leu* | 2.39 |
| 苏氨酸 Thr* | 1.65 | 酪氨酸 Tyr | 1.60 |
| 丝氨酸 Ser | 1.44 | 苯丙氨酸 Phe* | 1.27 |
| 谷氨酸 Glu | 5.80 | 赖氨酸 Lys* | 2.58 |
| 脯氨酸 Pro | 1.99 | 组氨酸 His* | 0.88 |
| 甘氨酸 Gly | 3.08 | 精氨酸 Arg | 2.30 |
| 丙氨酸 Ala | 2.46 | 色氨酸Trp* | 0.23 |
| 缬氨酸 Val* | 1.69 | 氨基酸总量TAA | 35.29 |
| 蛋氨酸 Met* | 0.60 | 必需氨基酸含量EAA | 12.86 |
| 异亮氨酸 Ile* | 1.57 | EAA/TAA | 36.44 |
注: *为必需氨基酸 |
表4 OYH联合5-FU对小鼠体重、瘤重和肿瘤抑制率的影响($\overline{x}$±SD, n=10)Tab. 4 Effects of OYH combined with 5-FU on body weight, tumor weight and tumor inhibition ratio of mice ($\overline{x}$ ± SD, n = 10) |
| 组 别 | 初体重/g | 终体重/g | 瘤重/g | 抑瘤率/% |
|---|---|---|---|---|
| 正常组 | 25.99±1.23 | 29.11±1.50 | — | — |
| 模型组 | 24.27±1.76 | 28.72±2.51 | 1.57±0.30 | — |
| 5-FU组 | 24.14±0.82 | 25.15±1.41***△△△ | 1.09±0.16*** | 31.00 |
| OYH-L+5-FU组 | 24.58±1.41 | 25.95±1.45**△△△ | 0.97±0.18*** | 38.24 |
| OYH-M+5-FU组 | 24.61±2.13 | 26.64±1.65*#△△ | 0.85±0.09***### | 46.27 |
| OYH-H+5-FU组 | 23.94±1.92 | 26.84±1.83#△△ | 0.82±0.19***## | 47.57 |
注: 与模型组比较: *表示p<0.05、**表示p<0.01、***表示p<0.001; 与5-FU组比较: #表示p<0.05、##表示p<0.01、###表示p<0.001; 与正常组比较: △表示p<0.05、△△表示p<0.01、△△△表示p<0.001 |
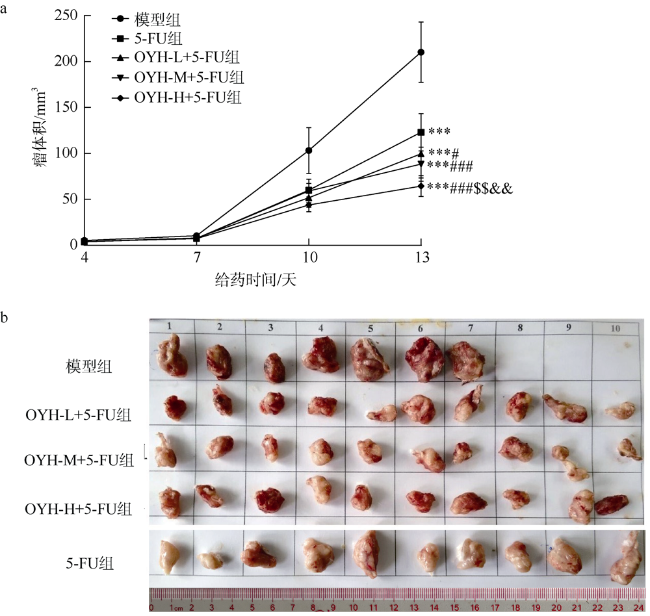
图2 OYH联合5-FU对S180荷瘤小鼠肿瘤体积大小的变化情况($\overline{x}$±SD, n=10)(a)和肿瘤解剖图(b)图a中***表示p<0.001 (与模型组比较); #表示p<0.05、###表示p<0.001 (与5-FU组比较); $$表示p<0.01 (与OYH-L+5-FU组比较); &&表示p<0.01 (与OYH-M+5-FU组比较) Fig. 2 Tumor volume of OYH combined with 5-FU on S180 tumor-bearing mice ($\overline{x}$ ± SD, n = 10) (a) and anatomical map of the tumor (b) |
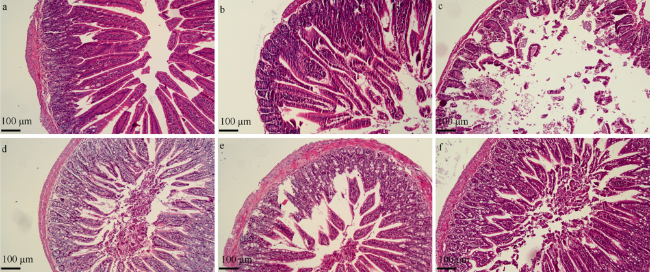
图3 OYH联合5-FU对小肠组织病理学切片染色图(10×10倍)a. 正常组; b. 模型组; c. 5-FU组; d. OYH-L+5-FU组; e. OYH-M+5-FU组; f. OYH-H+5-FU组 Fig. 3 OYH combined with 5-FU on histopathological staining of the small intestine (magnification10x). (a) Normal group; (b) Model group; (c) 5-FU group; (d) OYH-L+5-FU group; (e) OYH-M+5-FU group; (f) OYH-H+5-FU group |
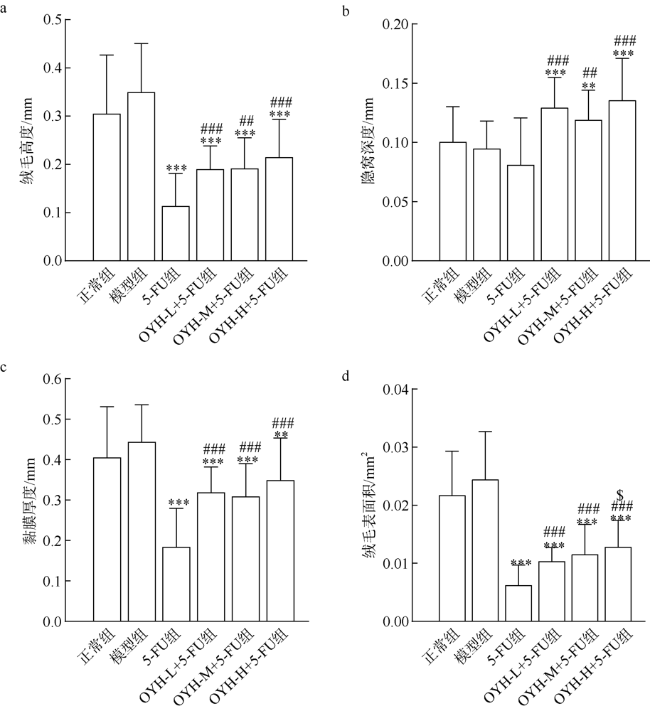
图4 OYH联合5-FU对小肠组织的形态学分析($\overline{x}$±SD, n=18)a. 绒毛高度; b. 隐窝深度; c. 黏膜厚度; d. 绒毛表面积。图中**表示p<0.01、***表示p<0.001 (与模型组比较); ##表示p<0.01、###表示p<0.001 (与5-FU组比较); $表示p<0.05 (与OYH-L+5-FU组比较) Fig. 4 OYH combined with 5-FU on histomorphological analyses of the small intestine ($\overline{x}$±SD, n = 18). (a) villus height; (b) crypt depth; (c) mucosa thickness; (d) villus surface area |
表5 OYH联合5-FU对小鼠血常规的影响($\overline{x}$±SD, n=10)Tab. 5 Effects of OYH combined with 5-FU on blood routine of mice ($\overline{x}$ ± SD, n = 10) |
| 正常组 | 模型组 | 5-FU组 | OYH-L+5-FU组 | OYH-M+5-FU组 | OYH-H+5-FU组 | |
|---|---|---|---|---|---|---|
| 白细胞/(×109L-1) | 6.61±2.58*** | 13.22±3.23 | 4.71±0.99*** | 4.25±1.14*** | 4.12±0.46*** | 4.40±0.81*** |
| 红细胞/(×1012L-1) | 10.34±0.46*** | 9.03±0.37 | 8.82±0.44 | 9.12±0.41 | 8.70±0.35$ | 8.72±0.49 |
| 血红蛋白/(g·L-1) | 158.90±6.79*** | 136.20±6.16 | 134.70±5.56 | 138.80±5.12 | 133.00±5.42$ | 133.10±7.46 |
| 红细胞压积/% | 44.93±1.68*** | 39.13±1.27 | 38.53±1.60 | 39.25±1.40 | 37.73±1.51*$ | 37.68±1.99 |
| 血小板/(×109L-1) | 1312.30±149.95*** | 1740.90±169.84 | 1601.50±196.78 | 1747.00±257.28 | 1640.40±170.42 | 1817.30±209.85# |
| 淋巴细胞/% | 80.29±4.17*** | 54.82±6.35 | 62.65±11.25 | 66.71±8.16** | 65.15±6.27** | 65.05±8.91** |
| 中性粒细胞/% | 17.47±4.08*** | 39.43±6.90 | 32.14±9.61 | 28.57±7.54** | 30.95±6.22** | 30.22±8.20* |
| 单核细胞/% | 0.87±0.40*** | 4.98±1.57 | 4.22±2.22 | 3.60±0.66* | 2.99±1.00** | 3.77±1.11 |
| 嗜酸性粒细胞/% | 1.34±0.45** | 0.74±0.19 | 0.99±0.51 | 1.10±0.46* | 0.91±0.48 | 0.89±0.57 |
| 嗜碱性粒细胞/% | 0.03±0.07 | 0.03±0.05 | 0.00±0.00 | 0.02±0.06 | 0.00±0.00 | 0.07±0.12 |
注: *表示p<0.05、**表示p<0.01、***表示p<0.001 (与模型组比较); #表示p<0.05、##表示p<0.01、###表示p<0.001 (与5-FU组比较); $表示p<0.05、$$表示p<0.01 (与OYH-L+5-FU组比较) |
表6 OYH联合5-FU对小鼠血清生化的影响($\overline{x}$±SD, n=10)Tab. 6 Effects of OYH combined with 5-FU on serum biochemical indicators of mice ($\overline{x}$±SD, n=10) |
| 正常组 | 模型组 | 5-FU组 | OYH-L+5-FU组 | OYH-M+5-FU组 | OYH-H+5-FU组 | |
|---|---|---|---|---|---|---|
| 总蛋白/(g·L-1) | 73.83±7.65*** | 57.35±7.11 | 54.72±6.55 | 61.39±6.85# | 64.36±8.95# | 70.56±8.85**###$ |
| 白蛋白/(g·L-1) | 48.41±4.62*** | 36.15±5.41 | 29.90±3.23** | 37.25±8.01# | 38.36±9.21# | 44.58±5.75**###$ |
| 前白蛋白/(mg·L-1) | 3.74±0.70** | 2.71±0.53 | 2.45±0.46 | 2.45±0.53 | 2.47±0.52 | 2.57±0.52 |
| 转铁蛋白/(g·L-1) | 10.31±1.54** | 8.52±1.08 | 8.64±1.21 | 9.25±1.06 | 9.42±1.18 | 9.32±1.00 |
| 补体C3 /(g·L-1) | 2.56±0.61*** | 3.78±0.41 | 3.09±1.12 | 3.70±0.52 | 3.84±0.79 | 3.56±0.88 |
| 补体C4 /(g·L-1) | 0.38±0.07*** | 0.53±0.06 | 0.40±0.08** | 0.43±0.09* | 0.43±0.07** | 0.43±0.08** |
| 免疫球蛋白A /(g·L-1) | 0.37±0.12*** | 0.13±0.03 | 0.12±0.03 | 0.15±0.03# | 0.17±0.03*## | 0.18±0.05*## |
| 免疫球蛋白G /(g·L-1) | 13.54±1.98** | 10.57±2.06 | 10.12±0.98 | 10.71±2.39 | 11.47±2.36 | 11.83±1.77# |
| 免疫球蛋白M /(g·L-1) | 1.72±0.33*** | 1.08±0.19 | 0.99±0.15 | 1.23±0.32# | 1.56±0.26***###$ | 1.64±0.25***### $$ |
注: *表示p<0.05、**表示p<0.01、***表示p<0.001 (与模型组比较); #表示p<0.05、##表示p<0.01、###表示p<0.001 (与5-FU组比较); $表示p<0.05、$$表示p<0.01 (与OYH-L+5-FU组比较) |
| [1] |
杜双双, 2018. 蚕丝蛋白肽免疫调节及与化疗的联合作用[D]. 天津: 天津医科大学:11-13.
|
| [2] |
胡雪琼, 吴红棉, 刘芷筠, 等, 2009. 近江牡蛎糖胺聚糖的酶解提取及其抗肿瘤活性研究[J]. 食品研究与开发, 30(7): 3-6.
|
| [3] |
胡雪琼, 吴红棉, 范秀萍, 等, 2014. 近江牡蛎糖胺聚糖的免疫调节活性研究[J]. 现代食品科技, 30(12): 16-24.
|
| [4] |
李婉, 曹文红, 章超桦, 等, 2017. 牡蛎酶解产物的组成特点及其体外免疫活性[J]. 食品工业科技, 38(16): 35-42.
|
| [5] |
巫楚君, 潘剑宇, 蔡冰娜, 等, 2021. 黄鳍金枪鱼酶解物免疫活性及其氨基酸分析[J]. 热带海洋学报, 40(6): 128-134.
|
| [6] |
武美彤, 张海欣, 张梦, 等, 2020. 牡蛎酶解物对Lewis肺癌的抑制作用及机制[J]. 食品与发酵工业, 46(11): 98-104, 111.
|
| [7] |
吴园涛, 任小波, 孙恢礼, 2012. 关于我国海洋生物资源领域科技布局的若干思考[J]. 热带海洋学报, 31(1): 79-84.
|
| [8] |
许东晖, 许实波, 王兵, 等, 1999. 皱纹盘鲍多糖抗肿瘤药理作用研究[J]. 热带海洋学报, 18(4): 86-90.
|
| [9] |
姚望, 张鑫杰, 姚庆华, 2019. 胃肠道恶性肿瘤化疗患者早期肠内营养干预的临床研究[J]. 中国肿瘤临床, 46(15): 780-784.
|
| [10] |
赵强, 魏祥玲, 孙建安, 等, 2021. 牡蛎资源的综合开发利用研究进展[J]. 中国食品添加剂, 32(7): 150-159.
|
| [11] |
中华人民共和国国家卫生和计划生育委员会, 国家食品药品监督管理总局, 2016. 食品安全国家标准食品中氨基酸的测定 GB 5009.6-2016[S]. 北京: 中国标准出版社.
NATIONAL HEALTH AND FAMILY PLANNING COMMISSION OF THE PEOPLE’S REPUBLIC OF CHINA, STATE FOOD AND DRUG ADMINISTRATION, 2016. National Food Safety Standards Determination of Amino Acids in Foods GB 5009.124-2016[S]. Beijing: Standards Press of China. (in Chinese with English abstract)
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}