
Journal of Tropical Oceanography >
Mitogenome characteristics and phylogenetic analysis of Siphonosoma australe in Hainan
Copy editor: YIN Bo
Received date: 2022-04-19
Revised date: 2022-07-22
Online published: 2022-07-27
Supported by
Hainan Provincial Natural Science Foundation of China(319MS013)
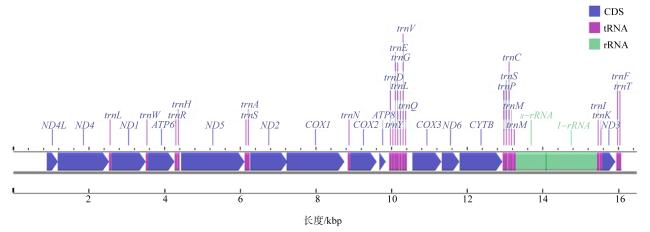
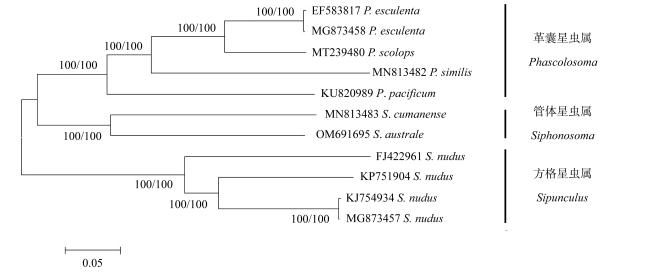
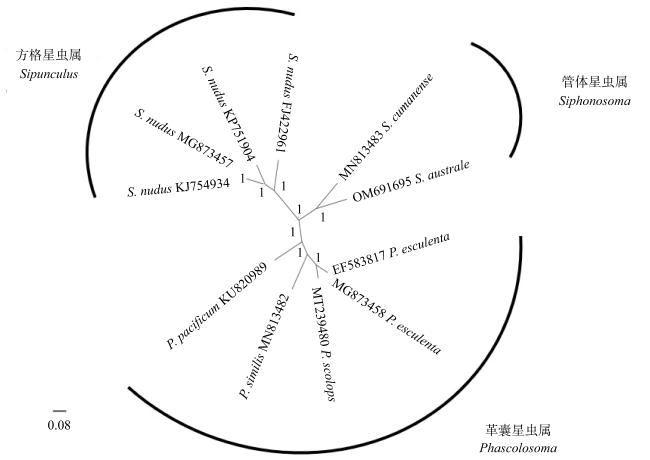
Siphonosoma australe is a local fishery resource in Hainan, which belongs to Siphonosoma. The taxonomic status of the genus in the phylum Sipuncula has been highly controversial. In this study, the mitogenome of S. australe in the Wenchang coast of Hainan was determined by high-throughput sequencing, and compared with the mitochondrial genomes collected from GenBank, characteristics of mitogenome sequence was analyzed, furthermore, the evolutionary position of the genus Siphonosoma in the phylum of Sipuncula was explored. The results show that the mitogenome of S. australe has 16483 base pairs and encodes a set of 38 genes (13 protein-coding, 23 transfer RNAs, and 2 ribosomal RNAs). The mitochondrial genome shows AT bias, and the content of nucleotides A+T is 65.87%. By the analysis of the relative synonymous codon usage (RSCU), the mitochondrial protein-coding genes of S. australe have obvious preference to the codons ending with A and U. The number of amino acids of mitochondrial protein coding genes (COX1, COX3 and ND5) of S. australe is quite different from that of other species. Compared with the mitochondrial genomes of sipunculans from GenBank, it was found that among the major coding genes of sipunculans, the proportions of variable sites in COX1, COX2 and CYTB genes are low, while the proportions of variable sites in ND2, ND4L and ND6 genes are high, and the highest proportion of variable sites in ATP8 gene is 83.33%. Based on the DNA sequences of sipunculan mitogenomes, the phylogenetic tree constructed using the methods of NJ (neighbor-joining), ML (maximum likelihood) and BI (Bayesian inference) showed that the genus Siphonosoma, as an independent clade, is between the genera of Phascolosoma and Sipunculus, even close to the genus of Phascolosoma instead far from the genus of Sipunculus. This result is inconsistent with the previously traditional classification based on morphological analyses. Taxonomically, Siphonosoma mixes the morphological characteristics of Phascolosoma and Sipunculus. The phylogenetic tree constructed with mitochondrial genome sequences could truly reflect the unique phylogenetic status of Siphonosoma. The mitogenome data of S. australe provides a theoretical basis for explaining the evolutionary relationship of the Siphonosoma in the phylum Sipuncula, and are beneficial to the protection and development of genetic resource of S. australe.
Key words: Siphonosoma australe; Hainan; mitogenome; phylogeny
HUANG Peixian , YAO Xuemei , YU Qiaochi , ZHANG Jiayu . Mitogenome characteristics and phylogenetic analysis of Siphonosoma australe in Hainan[J]. Journal of Tropical Oceanography, 2023 , 42(2) : 54 -63 . DOI: 10.11978/2022083
表1 外源信息表Tab. 1 List of the outer groups |
| 属 | 物种 | 采样地 | GenBank 登录号 |
|---|---|---|---|
| 方格星虫属Sipunculus | 光裸方格星虫(Sipunculus nudus) | Concarneau, France | FJ422961 |
| Beibu Bay, China | MG873457 | ||
| Gulei, China | KJ754934 | ||
| Yantai, China | KP751904 | ||
| 革囊星虫属Phascolosoma | 可口革囊星虫(Phascolosoma esculenta) | Beibu Bay, China | MG873458 |
| Wenzhou, China | EF583817 | ||
| 太平洋革囊星虫(Phascolosoma pacificum) | Chuuk, Micronesia | KU820989 | |
| 厥目革囊星虫(Phascolosoma scolops) | Beibu Bay, China | MT239480 | |
| 类革囊星虫(Phascolosoma similis) | Beibu Bay, China | MN813482 | |
| 管体星虫属Siphonosoma | 库岛管体星虫(Siphonosoma cumanense) | Beibu Bay, China | MN813483 |
表2 澳洲管体星虫线粒体基因组碱基组成Tab. 2 Composition of the mitochondrial genome in Siphonosoma australe |
| 基因序列 | T/% | C/% | A/% | G/% | (A+T)/% | (C+G)/% |
|---|---|---|---|---|---|---|
| 全基因组 | 36.54 | 20.97 | 29.33 | 13.16 | 65.87 | 34.13 |
| 蛋白质编码基因 | 37.49 | 21.56 | 27.86 | 13.09 | 65.35 | 34.65 |
| tRNA基因 | 31.04 | 18.02 | 32.79 | 18.15 | 63.83 | 36.17 |
| rRNA基因 | 33.01 | 19.12 | 32.46 | 15.41 | 65.47 | 34.53 |
表3 澳洲管体星虫线粒体蛋白质编码基因的相对同义密码子使用度(RSCU)Tab. 3 RSCU of mtDNA protein-coding sequence in Siphonosoma australe |
| 氨基酸 | 密码子 | 次数 | RSCU | 氨基酸 | 密码子 | 次数 | RSCU |
|---|---|---|---|---|---|---|---|
| Phe | UUU | 236 | 1.45 | Tyr | UAU | 91 | 1.35 |
| UUC | 90 | 0.55 | UAC | 44 | 0.65 | ||
| Leu | UUA | 226 | 2.17 | His | CAU | 56 | 1.30 |
| UUG | 20 | 0.19 | CAC | 30 | 0.70 | ||
| CUU | 173 | 1.66 | Gln | CAA | 62 | 1.70 | |
| CUC | 68 | 0.65 | CAG | 11 | 0.30 | ||
| CUA | 127 | 1.22 | Asn | AAU | 71 | 1.31 | |
| CUG | 10 | 0.10 | AAC | 37 | 0.69 | ||
| Ile | AUU | 226 | 1.52 | Lys | AAA | 79 | 1.82 |
| AUC | 71 | 0.48 | AAG | 8 | 0.18 | ||
| Met | AUA | 161 | 1.71 | Asp | GAU | 34 | 1.05 |
| AUG | 27 | 0.29 | GAC | 31 | 0.95 | ||
| Val | GUU | 78 | 1.70 | Glu | GAA | 63 | 1.70 |
| GUC | 25 | 0.55 | GAG | 11 | 0.30 | ||
| GUA | 73 | 1.60 | Cys | UGU | 24 | 1.33 | |
| GUG | 7 | 0.15 | UGC | 12 | 0.67 | ||
| Ser | UCU | 117 | 2.77 | Trp | UGA | 87 | 1.81 |
| UCC | 41 | 0.97 | UGG | 9 | 0.19 | ||
| UCA | 84 | 1.99 | Arg | CGU | 22 | 1.22 | |
| UCG | 7 | 0.17 | CGC | 11 | 0.61 | ||
| AGU | 19 | 0.45 | CGA | 33 | 1.83 | ||
| AGC | 13 | 0.31 | CGG | 6 | 0.33 | ||
| AGA | 51 | 1.21 | Pro | CCU | 71 | 1.41 | |
| AGG | 6 | 0.14 | CCC | 29 | 0.57 | ||
| Thr | ACU | 93 | 1.52 | CCA | 95 | 1.88 | |
| ACC | 38 | 0.62 | CCG | 7 | 0.14 | ||
| ACA | 102 | 1.67 | Gly | GGU | 44 | 0.91 | |
| ACG | 12 | 0.20 | GGC | 28 | 0.58 | ||
| Ala | GCU | 96 | 1.39 | GGA | 98 | 2.02 | |
| GCC | 53 | 0.77 | GGG | 24 | 0.49 | ||
| GCA | 115 | 1.67 | |||||
| GCG | 12 | 0.17 |
表4 星虫动物3个属7个种的线粒体蛋白质编码基因的氨基酸数量Tab. 4 Amino acid quantity of protein-coding genes in sipunculan mitochondrial genomes of 7 species (3 genera) |
| 星虫名称 | ATP6 | ATP8 | CYTB | COX1 | COX2 | COX3 | ND1 | ND2 | ND3 | ND4 | ND4L | ND5 | ND6 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 可口革囊星虫 | 231 | 54 | 378 | 519 | 231 | 267 | 304 | 323 318 | 121 | 450 451 | 94 | 571 562 | 157 |
| 太平洋革囊星虫 | 231 | 49 | 378 | 518 | 227 | 263 | 308 | 319 | 118 | 450 | 94 | 572 | 157 |
| 类革囊星虫 | 231 | 53 | 378 | 519 | 230 | 267 | 303 | 327 | 121 | 450 | 94 | 571 | 157 |
| 厥目革囊星虫 | 231 | 54 | 378 | 519 | 229 | 291 | 304 | 320 | 121 | 451 | 94 | 572 | 157 |
| 光裸方格星虫 | 229 234 | 52 53 | 379 | 516 519 | 231 230 | 259 | 314 312 | 328 327 329 | 119 117 | 452 454 | 93 | 565 571 | 157 |
| 库岛管体星虫 | 231 | 54 | 379 | 513 | 231 | 275 | 303 | 318 | 114 | 448 | 94 | 569 | 155 |
| 澳洲管体星虫 | 231 | 56 | 377 | 510 | 232 | 293 | 303 | 327 | 116 | 452 | 94 | 559 | 155 |
注: 同一栏目中不同的数字表示同一物种的不同线粒体蛋白质编码基因的氨基酸数量差异 |
表5 星虫动物3个属7个种的线粒体基因组的主编码基因变异位点分析Tab. 5 Genetic variation analysis of major encoding genes in sipunculan mitochondrial genomes of 7 species (3 genera) |
| 基因名称 | 总位点数 | 保守位点数 | 变异位点数 | 变异位点比例/% |
|---|---|---|---|---|
| ATP6 | 669 | 226 | 443 | 66.22 |
| ATP8 | 144 | 24 | 120 | 83.33 |
| CYTB | 1131 | 550 | 581 | 51.37 |
| COX1 | 1531 | 824 | 707 | 46.18 |
| COX2 | 680 | 328 | 352 | 51.76 |
| COX3 | 780 | 342 | 438 | 56.15 |
| ND1 | 906 | 368 | 538 | 59.38 |
| ND2 | 944 | 215 | 729 | 77.22 |
| ND3 | 344 | 126 | 218 | 63.37 |
| ND4 | 1329 | 402 | 927 | 69.75 |
| ND4L | 282 | 77 | 205 | 72.70 |
| ND5 | 1657 | 523 | 1134 | 68.44 |
| ND6 | 466 | 118 | 348 | 74.68 |
| 12S rRNA | 753 | 295 | 458 | 60.82 |
| 16S rRNA | 980 | 423 | 557 | 56.84 |
| [1] |
陈子安, 杜晓东, 王庆恒, 等, 2007. 3种星虫线粒体16S rRNA、COI和Cytb基因片段的序列比较[J]. 广东海洋大学学报, 27(4): 3-10.
|
| [2] |
傅静, 孙啸, 2003. 基于全基因组的系统发生分析[J]. 生物技术, (6): 53-56.
|
| [3] |
黄族豪, 刘迺发, 2010. 动物线粒体基因组变异研究进展[J]. 生命科学研究, 14(2): 166-171.
|
| [4] |
蒋文枰, 李家乐, 郑润玲, 等, 2010. 褶纹冠蚌线粒体基因组全序列分析[J]. 遗传, 32(2): 153-162.
|
| [5] |
兰国宝, 杨素芳, 谢体三, 等, 2007. 星虫动物门系统发生研究进展[J]. 广西科学, 14(2): 186-192.
|
| [6] |
李凤鲁, 周红, 王玮, 1992. 中国沿海管体星虫属(星虫动物门)的研究[J]. 青岛海洋大学学报: 自然科学版, 22(1): 97-102.
|
| [7] |
孟乾, 张志勇, 张志伟, 等, 2020. 斑石鲷和条石鲷线粒体基因组密码子使用分析[J]. 水产科学, 39(5): 702-709.
|
| [8] |
彭银辉, 周于娜, 刘旭佳, 等, 2017. 基于线粒体控制区序列的光裸方格星虫群体遗传多样性分析[J]. 水产学报, 41(10): 1542-1551.
|
| [9] |
乔立君, 姚雪梅, 余巧驰, 2022. 海南澳洲管体星虫卵细胞发育及生殖周期研究[J]. 热带海洋学报, 41(5): 161-169.
|
| [10] |
申欣, 2012. 星虫动物线粒体基因组全序列的基因排列和特征比较[J]. 水产科学, 31(9): 554-559.
|
| [11] |
宋素霞, 2015. 中国沿海光裸方格星虫线粒体基因组分析及遗传多样性研究[D]. 福建: 集美大学.
|
| [12] |
宋素霞, 丁少雄, 鄢庆枇, 等, 2017. 中国沿海光裸方格星虫6个地理群体遗传多样性分析[J]. 水产学报, 41(8): 1183-1191.
|
| [13] |
孙萌, 2021. 基于多基因对寡膜纲纤毛虫重要类群分子系统学的研究[D]. 黑龙江: 哈尔滨师范大学.
|
| [14] |
夏立萍, 徐鸣, 郭宝英, 等, 2021. 等边浅蛤线粒体全基因组测序和系统发育研究[J]. 浙江海洋大学学报(自然科学版), 40(2): 93-100.
|
| [15] |
于黎, 张亚平, 2006. 系统发育基因组学——重建生命之树的一条迷人途径[J]. 遗传, 28(11): 1445-1450.
|
| [16] |
钟声平, 蒋艳, 刘永宏, 等, 2020. 广西5种常见星虫动物的线粒体基因组特征和系统进化分析[J]. 广西科学, 27(5): 532-540.
|
| [17] |
周红, 李凤鲁, 王玮, 2007. 中国动物志, 无脊椎动物. 第四十六卷, 星虫动物门螠虫动物门[M]. 北京: 科学出版社.
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}