
Journal of Tropical Oceanography >
Difference in gene expression of Aiptasia pulchella after albino recovery
Copy editor: LIN Qiang
Received date: 2020-12-04
Revised date: 2021-02-24
Online published: 2021-02-25
Supported by
Fujian Province Young and Middle-aged Teacher Education Research Project(JT180386)
Fujian Natural Science Foundation Project(2020J01855)
Copyright
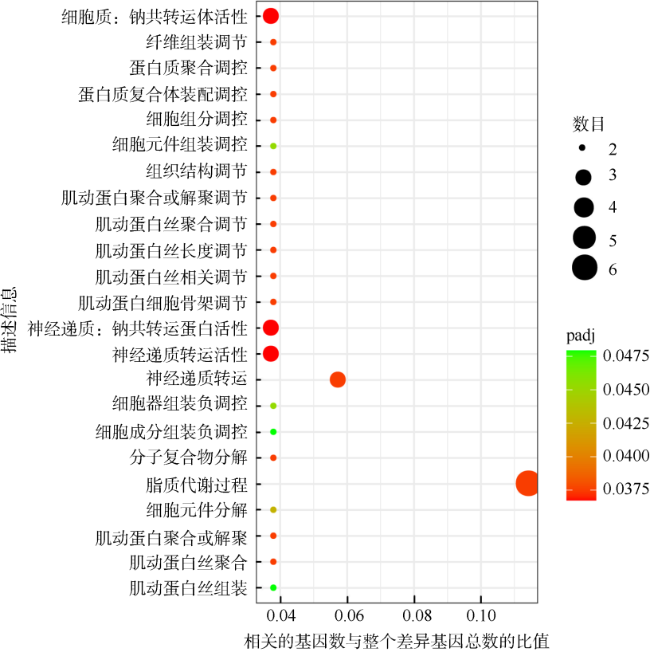
Anemone bleaching can be defined generally as a decline in the population of the symbiotic algae, a reduction in the pigment content of the algae, or both. In order to study the molecular mechanism of Aiptasia pulchella during bleaching and recovered phage affected by chronic heat stress, transcriptome analysis by Illumina Hi-seq of bleaching and recovered A. pulchella was carried out on 50109686 and 43163786 clean reads, which were annotated as 24565 and 24157 unigenes, respectively. The results of comparative transcriptome analysis showed that there were 214 differentially expressed genes (DEGs), including 101 that were significantly highly expressed in bleaching A. pulchella and 113 in recovered A. pulchella. These genes were mainly involved in DNA replication, metabolism, ion transport, and collagen. Gene set enrichment analysis (GSEA) of all 28, 050 expressed genes showed that collagen and ion transport were enriched in bleaching A. pulchella, while nucleotide repair was enriched in recovered A. pulchella. We speculate that these biological processes may play an important role in the bleaching and recovering phases of anemone. The main findings obtained in this study preliminarily reveal the molecular mechanism of adaptive regulation of anemone holobiont after symbiotic imbalance, providing a theoretical basis for the adaptive mechanism of anemone and coral symbiosis system in response to environmental changes.
Key words: Aiptasia pulchella; Transcriptome; Bleaching; Recovery after bleaching
ZHANG Qian , YAN Hao , WANG Lu . Difference in gene expression of Aiptasia pulchella after albino recovery[J]. Journal of Tropical Oceanography, 2021 , 40(6) : 111 -119 . DOI: 10.11978/2020143
表1 鬼手海葵的转录组测序基本信息Tab. 1 Information of transcriptome sequencing of A. pulchella |
| 样品名称 | Raw reads | Clean reads | Clean bases | Error rate | Q20/% | Q30/% | GC pct/% | Gene number | Novel |
|---|---|---|---|---|---|---|---|---|---|
| AP_W | 51938216 | 50109686 | 7.52G | 0.03 | 96.52 | 90.82 | 41.42 | 24565 | 1886 |
| AP_H | 43920530 | 43163786 | 6.47G | 0.03 | 97.86 | 93.97 | 43.42 | 24157 | 1869 |
注: Raw reads, 原始数据中的reads数; Clean reads, 原始数据过滤后的reads数; Clean bases, 原始数据过滤后的碱基数; Error rate, 数据整体测序错误率; Q20, Phred数值大于20的碱基占总碱基的百分比; Q30, Phred数值大于30的碱基占总碱基的百分比; GC pct, clean reads中G与C占四种碱基的百分比; Gene number, 基因数目; Novel, 新基因数目 |
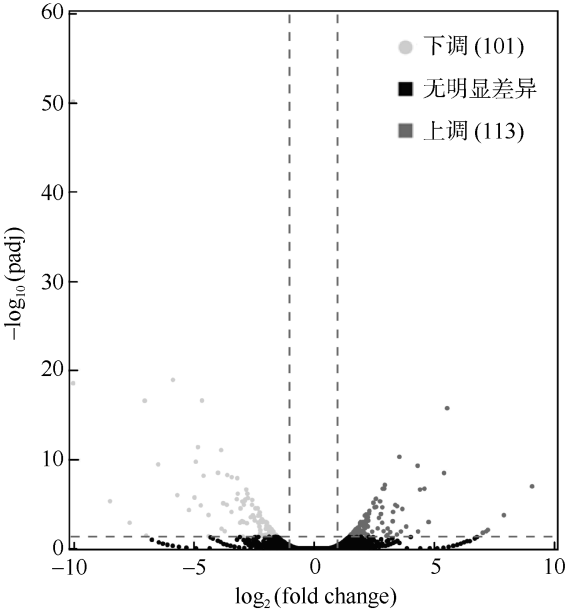
图2 白化海葵和恢复的海葵差异表达基因火山图浅灰色表示下调, 即白化恢复的海葵显著低表达基因有101个; 深灰色表示上调, 即白化恢复的海葵显著高表达的基因有113个 ; 黑色表示无显著差异; -log10(padj)为基因差异表达显著性; |log2(fold change)|表示基因表达量差异倍数 Fig. 2 Volcano plot of differentially expressed genes between bleaching and recovering A. pulchella. Light gray indicates 101 down-regulated in recovering A. pulchella, and dark grey indicates 113 up-regulated in recovering A. pulchella. Black indicates no significant difference. -log10(padj) is significant difference in gene expression. |log2(fold change)| indicates the fold change of gene expression. |
表2 与DNA复制、代谢、离子转运和胶原蛋白相关的差异表达基因Tab. 2 Differentially expressed genes associated with DNA replication, metabolism, ion transport, and collagen |
| 基因功能 | 基因编号 | AP_W | AP_H | padj | 基因注释 |
|---|---|---|---|---|---|
| 与DNA复制相关基因 | gene25952 | 1.69 | 37.61 | <0.01 | ATP-dependent DNA helicase PIF1 |
| gene16761 | 8.50 | 64.63 | <0.01 | ATP-dependent DNA helicase PIF1 | |
| gene20928 | 52.76 | 172.71 | 0.04 | ATP-dependent DNA helicase PIF1 | |
| gene1065 | 0.84 | 85.78 | <0.01 | Histone H2B type 1 | |
| novel.1702 | 27.23 | 152.74 | <0.01 | DDE superfamily endonuclease | |
| 与新陈代谢相关的基因 | gene9753 | 517.51 | 125.72 | <0.01 | Group XV phospholipase A2 |
| gene4747 | 52.76 | 760.13 | <0.01 | Beta-glucuronidase | |
| gene21985 | 68.93 | 9.41 | <0.01 | Beta-glucuronidase | |
| gene4924 | 1469.14 | 187.99 | <0.01 | Carbonic anhydrase 2 | |
| gene5393 | 204.27 | 748.38 | <0.01 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 9 | |
| gene12810 | 634.97 | 1875.04 | 0.03 | Acetyl-CoA carboxylase | |
| gene22575 | 32.33 | 157.44 | <0.01 | Aldehyde dehydrogenase family 3 member B1 | |
| gene22820 | 148.95 | 37.61 | 0.01 | Chitobiase | |
| gene24483 | 52.76 | 180.94 | 0.03 | D-3-phosphoglycerate dehydrogenase | |
| gene9970 | 659.66 | 132.77 | <0.01 | Endoglucanase E | |
| gene898 | 11.90 | 189.16 | <0.01 | Lathosterol oxidase | |
| gene20346 | 676.68 | 2193.42 | 0.01 | Phosphatidylserine decarboxylase proenzyme | |
| 与离子转运有关的蛋白 | gene4543 | 128.52 | 566.28 | <0.01 | Ammonium transporter Rh type B |
| gene4000 | 520.06 | 170.36 | 0.03 | Voltage-dependent N-type calcium channel subunit alpha-1B | |
| gene1518 | 187.25 | 56.41 | 0.04 | Organic cation transporter protein | |
| gene10353 | 28.08 | 104.57 | 0.03 | Sodium-dependent neutral amino acid transporter B(0)AT3 | |
| gene13262 | 780.53 | 2478.91 | 0.01 | Reticulocalbin-3 | |
| gene23383 | 0.00 | 259.65 | <0.01 | Sodium- and chloride-dependent GABA transporter 3 | |
| gene10062 | 236.62 | 65.80 | 0.01 | 2-oxoglutarate and iron-dependent oxygenase domain-containing protein 2 | |
| 与胶原蛋白有关的基因 | novel.1027 | 684.34 | 180.94 | <0.01 | Collagen triple helix repeat |
| gene24960 | 18.71 | 0.00 | 0.01 | Tenascin-R | |
| gene2049 | 114.05 | 353.64 | 0.03 | Matrilin-2 | |
| gene19290 | 394.09 | 82.25 | <0.01 | Collagen alpha-1(X) chain | |
| gene18911 | 868.20 | 112.80 | <0.01 | Collagen alpha-5(Ⅵ) chain | |
| gene18914 | 432.39 | 74.03 | <0.01 | Collagen alpha-6(Ⅵ) chain | |
| gene11395 | 30.63 | 2.36 | 0.01 | Collagen triple helix repeat-containing protein 1 | |
| gene1876 | 421.33 | 91.65 | <0.01 | Fibrinogen C domain-containing protein 1 | |
| gene20101 | 490.27 | 156.27 | 0.03 | Fibulin-1 | |
| gene19080 | 117.45 | 28.21 | 0.01 | Gelsolin-like protein 1 | |
| gene8325 | 62.12 | 12.94 | 0.03 | Short-chain collagen C4 |
注: padj即经过FDR校正的P值(adjust p-value) |
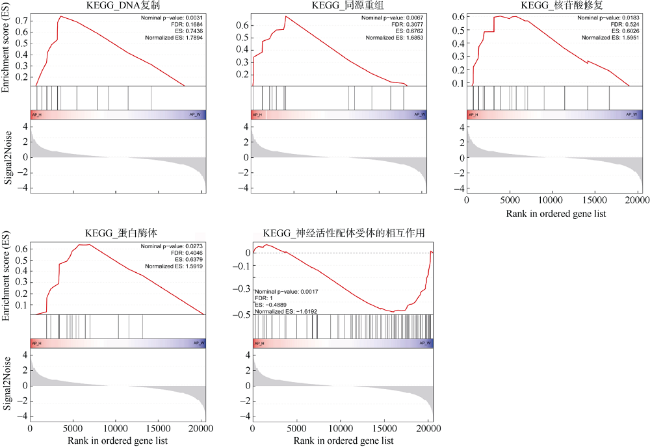
图4 GSEA富集中差异显著的KEGG基因集(Nominal p-value<0.05)FDR: 错误发现率; Enrichment score (ES): 富集评分; Normalized ES: 归一化的富集评分; Signal2Noise: 信噪比; Ranked in ordered gene set: 基因在特定基因集中的分布 Fig. 4 Significantly different KEGG pathway of Gene Set Enrichment Analysis (Nominal p-value<0.05). FDR indicates false discovery rate. ES indicates enrichment score. Normalized ES indicates normalized enrichment score. Signal2Noise indicates signal to noise ratio; Ranked in ordered gene set, the distribution of the gene ranked in ordered gene set. |
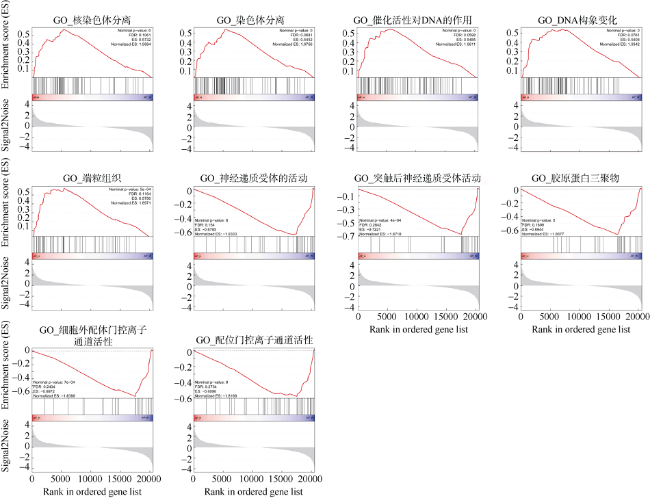
图5 GSEA富集中差异显著的GO基因集(Nominal p-value<0.05)FDR: 错误发现率; ES: 富集评分; Normalized ES: 归一化的富集评分; Signal2Noise:信噪比; Ranked in ordered gene set: 基因在特定基因集中的分布 Fig. 5 Significantly different GO pathway of Gene Set Enrichment Analysis (Nominal p-value<0.05). FDR indicates false discovery rate. ES indicates enrichment score. Normalized ES indicates normalized enrichment score. Signal2Noise indicates signal to noise ratio; Ranked in ordered gene set, the distribution of the gene ranked in ordered gene set. |
| [1] |
黄晖, 许昌有, 袁涛, 2013. 造礁石珊瑚白化相关功能基因的研究进展[J]. 热带海洋学报, 32(4):43-50.
|
| [2] |
李新正, 寇琦, 王金宝, 等, 2020. 中国海洋无脊椎动物分类学与系统演化研究进展与展望[J]. 海洋科学, 44(7):26-70.
|
| [3] |
李秀保, 黄晖, 练健生, 等, 2007. 珊瑚及共生藻在白化过程中的适应机制研究进展[J]. 生态学报, 27(3):1217-1225.
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}