
Journal of Tropical Oceanography >
Molecular identification, secondary metabolites and biological activities of a deep-sea-derived fungus 101#*
Received date: 2022-11-11
Revised date: 2022-12-26
Online published: 2022-12-28
Supported by
National Natural Science Foundation of China(31902373)
Natural Science Foundation of Guangdong Province, China(2021A1515011398)
Project of Education Department of Guangdong Province, China(2021ZDJS034)
Project of Education Department of Guangdong Province, China(2021KCXTD039)
Project of Education Department of Guangdong Province, China(2022-K08)
Public Service Platform of Biomedical Resources Research and Development of South China Sea(2017C8B2)
Yanling Outstanding Young Teacher Training Program of Lingnan Normal University(YL20200213)
Open Project of Mangrove Research Institute, Lingnan Normal University(PYXM05)
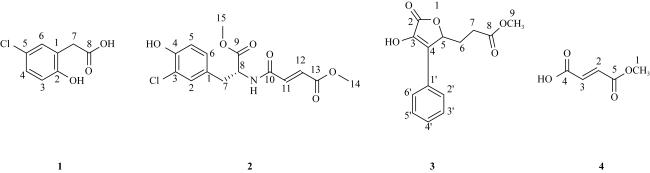
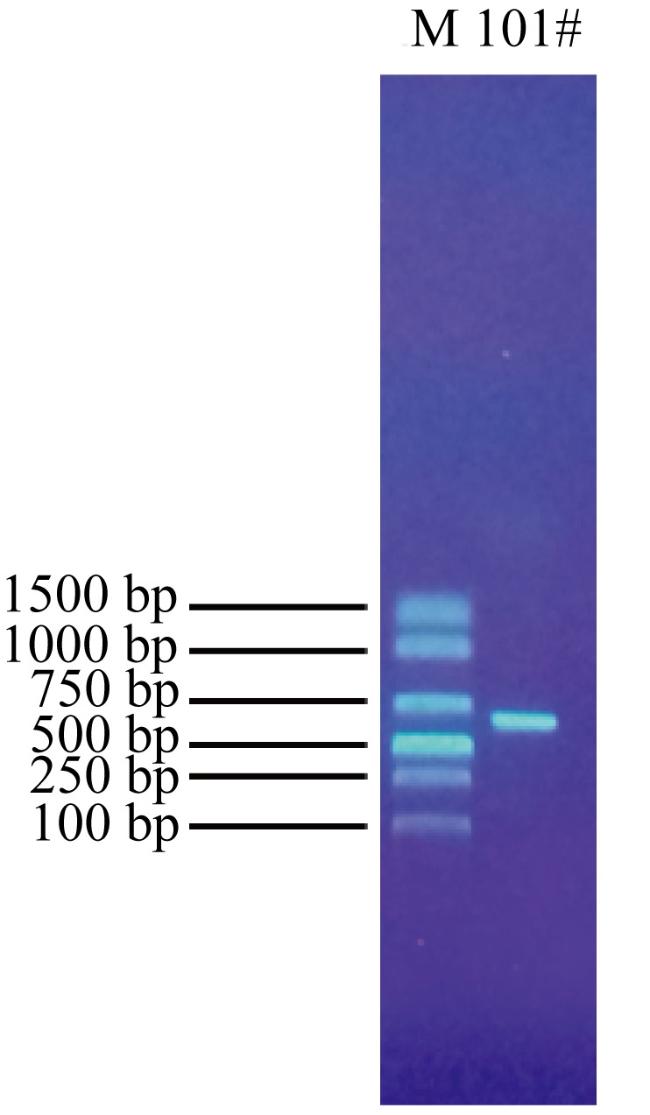
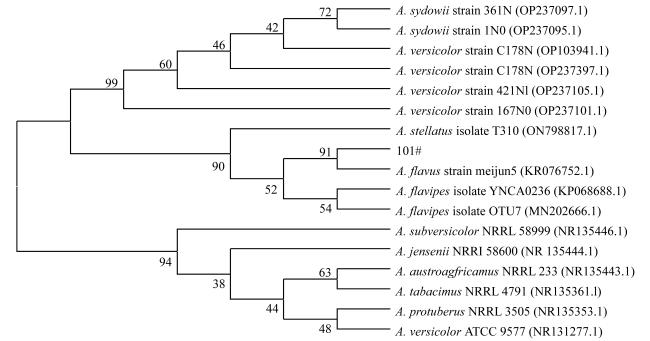
In order to provide reference basis for development and application of deep-sea microbial resources, the molecular identification, secondary metabolites and biological activity of a fungal strain (named as 101#) isolated from deep-sea sediment of the South China Sea was studied. The fungal strain was analyzed based on rDNA-ITS gene sequence. The changes in the content of its fermentation products were tracked by high performance liquid chromatography (HPLC). The secondary metabolites were isolated by silica gel column chromatography, Sephadex LH-20 and HPLC. The structures of compounds were identified by nuclear magnetic resonance (NMR), mass spectrometry (MS), and literature analysis. The biological activities were detected by filter paper-agar diffusion and brine shrimp lethal methods, respectively. The results show that the fungal strain 101# was identified as Aspergillus flavus. The optimal fermentation period was 28 d. In addition, the crude extract had certain inhibitory activities against 15 indicator bacteria. The minimum inhibitory concentration (MIC) against 10 Gram-positive and negative bacteria and 3 plant pathogenic bacteria were 0.781 ~ 6.25 μg·mL-1 and 50 ~ 200 μg·mL-1, respectively, and the MIC against marine biofouling bacterium (Alteromonas macleodii) and pathogenic fungus (Candida albicans) were 3.125 μg·mL-1 and 1.563 μg·mL-1, respectively. The crude extract had lethal activity against brine shrimp, and their LD50 at 1, 12, 24 and 48 h were 74.597, 24.322, 13.797 and 8.559 mg·mL-1, respectively. Finally, 4 monomer compounds were isolated from the ethyl acetate extraction site of the strain and their structures were identified as 5-chloro-2-hydroxyphenylacetic acid (1), aspergamide A (2), WF-3681 methyl Ester (3) and (E)-But-2-enedioic acid monomethyl Ester (4). The four compounds showed certain inhibitory activities against B. subtilis, C. albicans, B. thuringiensis, Shewanella and M. luteus, which MIC value were 6.25 ~ 200 μg·mL-1. Furthermore, compounds 1 ~ 4 showed strong brine shrimp lethal activity with LD50 values of 7.40, 11.95, 17.69, 23.35 μg·mL-1, respectively.
ZENG Buyan , LIANG Zhifeng , LUO Qinqin , ZENG Ling , YANG Changgeng , WANG Liyun , SUN Yulin . Molecular identification, secondary metabolites and biological activities of a deep-sea-derived fungus 101#*[J]. Journal of Tropical Oceanography, 2023 , 42(5) : 104 -114 . DOI: 10.11978/2022242
表1 菌株101#粗提物对12种指示菌的抑制活性及最小抑制浓度(MIC值)(n=3)Tab. 1 Minimum inhibitory concentration (MIC) of crude extract of strain 101# against 12 indicator bacteria (n=3) |
| 指示菌 | 阳性对照 | 粗提物浓度/(μg·mL-1) | MIC/(μg·mL-1) | |||||
|---|---|---|---|---|---|---|---|---|
| 12.5 | 6.25 | 3.125 | 1.563 | 0.781 | 0.391 | |||
| 枯草芽孢杆菌 (Bacillus subtilis) | ++++ | ++ | ++ | + | + | + | - | 0.781 |
| 藤黄微球菌 (Micrococcus luteus) | ++++ | ++ | ++ | + | + | - | - | 1.563 |
| 苏云金芽孢杆菌 (Bacillus thuringiensis) | ++++ | ++ | + | + | + | - | - | 1.563 |
| 耐甲氧西林金黄色葡萄球菌 (Methicillin-resistant staphylococcus aureus) | ++++ | ++ | + | + | - | - | - | 3.125 |
| 金黄色葡萄球菌 (Staphylococcus aureus) | ++++ | ++ | + | + | - | - | - | 3.125 |
| 无乳链球菌 (Streptococcus agalactiae) | ++++ | ++ | + | - | - | - | - | 6.250 |
| 稀瓦氏菌 (Shewanella) | ++++ | ++ | + | + | + | - | - | 1.563 |
| 大肠杆菌 (Escherichia coli) | ++++ | ++ | + | + | - | - | - | 3.125 |
| 福氏志贺菌 (Shigella Flexneri) | ++++ | ++ | + | + | - | - | - | 3.125 |
| 肺炎克雷伯菌 (Klebsiella pneumoniae) | ++++ | + | + | - | - | - | - | 6.250 |
| 麦氏交替单胞菌 (Alteromonas macleodii) SCAU-003 | ++++ | +++ | ++ | + | - | - | - | 3.125 |
| 多耐白色念珠菌 (Candida albicans) | ++++ | +++ | ++ | + | + | - | - | 1.563 |
注: -: 无抑制作用; +: 抑菌圈为6~9mm; + +: 抑菌圈为9~12mm; + + +: 抑菌圈为12~15mm; + + + +: 抑菌圈≥15mm。阳性对照组为硫酸庆大霉素 |
表2 菌株101#粗提物对6种植物病原菌的抑制活性及最小抑制浓度(MIC值)(n=3)Tab. 2 Minimum inhibitory concentration (MIC) of crude extract of strain 101# against 6 plant pathogens (n=3) |
| 指示菌 | 阳性对照 | 粗提物浓度/(μg·mL-1) | MIC/(μg·mL-1) | ||||
|---|---|---|---|---|---|---|---|
| 200 | 100 | 50 | 25 | 12.5 | |||
| 禾谷镰刀菌 (Fusarium graminearum) | ++ | ++ | + | + | - | - | 50 |
| 意大利青霉 (Penicillium italicum) | ++++ | + | - | - | - | - | 200 |
| 尖孢镰刀菌 (Fusarium oxysporum) | + | + | - | - | - | - | 200 |
| 层出镰刀菌 (Fusarium proliferatum) | + | - | - | - | - | - | >200 |
| 盘长孢状刺盘孢 (Colletotrichum gloeosprioides) | + | - | - | - | - | - | >200 |
| 茄镰孢菌 (Fusarium solani) | ++ | - | - | - | - | - | >200 |
注: -: 无抑制作用; +: 抑菌圈为6~9mm; + +: 抑菌圈为9~12mm; + + +: 抑菌圈为12~15mm; + + + +: 抑菌圈≥15mm。对照组为两性霉素B |
表3 菌株101#粗提物对卤虫致死活性的测试结果Tab 3. Test results of lethal activity of strain 101# crude extract against brine shrimp |
| 时间/h | 粗提物浓度/( mg·mL-1) | LD50/(mg·mL-1) | 95%置信区间 | |||||
|---|---|---|---|---|---|---|---|---|
| 100 | 50 | 25 | 12.5 | 6.25 | 下限 | 上限 | ||
| 1 | 64.7 | 38.9 | 25.0 | 10.0 | 0.0 | 74.597 | 54.900 | 118.483 |
| 12 | 100.0 | 83.3 | 56.3 | 30.0 | 12.5 | 24.322 | 17.238 | 34.071 |
| 24 | 100.0 | 100.0 | 87.5 | 35.5 | 18.8 | 13.797 | 9.739 | 19.393 |
| 48 | 100.0 | 100.0 | 93.8 | 65.0 | 37.5 | 8.559 | 4.888 | 12.455 |
表4 化合物1~4对5株指示菌的MIC值(单位: μg·mL-1)Tab. 4 Minimum inhibitory concentration (MIC) of compounds 1 ~ 4 against 5 indicator bacteria |
| 化合物 | 枯草芽孢杆菌 | 多耐白色念珠菌 | 苏云金芽孢杆菌 | 希瓦氏菌 | 藤黄微球菌 |
|---|---|---|---|---|---|
| 1 | 25 | 12.5 | 50 | 200 | >200 |
| 2 | >200 | 6.25 | 25 | >200 | 100 |
| 3 | 25 | 12.5 | 12.5 | >200 | >200 |
| 4 | 12.5 | 6.25 | 50 | 200 | 12.5 |
| 硫酸庆大霉素 | 12.5 | 6.25 | 1.56 | 12.5 | 12.5 |
*藤黄微球菌、无乳链球菌、福氏志贺菌、麦氏交替单胞菌以及6种植物病原菌均由华南农业大学张晓勇课题组惠赠, 在此表示衷心的感谢!
| [1] |
安昶亮, 2019. 海洋真菌Aspergillus sp. SCS-KFD66的次级代谢产物及其活性研究[D]. 南京: 南京农业大学:1-121.
|
| [2] |
董怡飞, 徐秀丽, 张新婉, 等, 2022. 深海真菌Aspergillus sp.20220129的次级代谢产物及抑菌活性研究[J]. 中国抗生素杂志, 47(5): 494-500.
|
| [3] |
黄智, 王发左, 田新朋, 等, 2012. 一株南海北部深海沉积环境真菌00457的鉴定及其活性研究[J]. 生物技术通报, (10): 199-204.
|
| [4] |
江北, 吕梦霞, 蒋冬花, 2019. 曲霉属真菌活性代谢产物及在农业生产中的应用研究进展[J]. 微生物学杂志, 39(2): 103-110.
|
| [5] |
罗寒, 李晓栋, 李晓明, 等, 2017. 红树林来源内生真菌杂色曲霉Aspergillus versicolor MA-229次级代谢产物研究[J]. 中国抗生素杂志, 42(4): 334-340.
|
| [6] |
潘力, 崔翠, 王斌, 2010. 一种用于PCR扩增的丝状真菌DNA快速提取方法[J]. 微生物学通报, 37(3): 450-453.
|
| [7] |
钱生辉, 2022. 南海红树林底泥海洋放线菌抗菌/抗群体感应活性菌株的筛选及次级代谢产物的初步研究[D]. 扬州: 扬州大学: 1-90.
|
| [8] |
单体壮, 2020. 海洋真菌Aspergillus ruber的次级代谢产物及其生物活性研究[D]. 扬州: 扬州大学: 1-137.
|
| [9] |
谭雁鸿, 李基兴, 林秀萍, 等, 2019. 中国南海软珊瑚真菌Eupenicillium sp. DX-SER3 (KC871024) 的次级代谢产物研究[J]. 热带海洋学报, 38(2): 43-47.
|
| [10] |
王发左, 2008. 海洋真菌抗肿瘤活性次级代谢产物及其生物转化研究[D]. 青岛: 中国海洋大学: 21-22.
|
| [11] |
叶禹秀, 罗小卫, 杨斌, 等, 2022. 南海礁栖海藻共附生真菌Pestalotiopsis neglecta SCSIO41 403次级代谢产物研究[J]. 热带海洋学报, 41(3): 186-190.
|
| [12] |
于清武, 胡丽琴, 李菲, 等, 2015. 南海深海沉积物可培养细菌多样性及其生物毒性分析[J]. 南方农业学报, 46(12): 2203-2208.
|
| [13] |
曾奇, 仲伟茂, 向瑶, 等, 2018. 南海深海沉积物中52株真菌的初步分离鉴定及其代谢产物活性[J]. 微生物学通报, 45(9): 1904-1915.
|
| [14] |
张长生, 李文利, 2018. 海洋微生物学: 新机遇, 新挑战[J]. 微生物学通报, 49(5): 1841-1842.
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}