
Journal of Tropical Oceanography >
Study on species composition and distribution of fish eggs in Yongle Atoll and Dongdao Island by high-throughput sequencing technology
Copy editor: LIN Qiang
Received date: 2022-12-28
Revised date: 2023-02-22
Online published: 2023-02-27
Supported by
Hainan Natural Science Foundation(322CXTD530)
Hainan Natural Science Foundation(322MS153)
Special Financial Project of Ministry of Agriculture and Rural Affairs(NFZX2021)
Basic and Applied Basic Research Project of Guangdong Province(2019B030302004-05)
Central Public-interest Scientific Institution Basal Research Fund, CAFS(2023TD16)
Special Fund Project for Basic Scientific Research Business Expenses of Central Public Welfare Scientific Research Institutes of South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences(2021SD04)
Special Fund Project for Basic Scientific Research Business Expenses of Central Public Welfare Scientific Research Institutes of South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences(2019TS28)
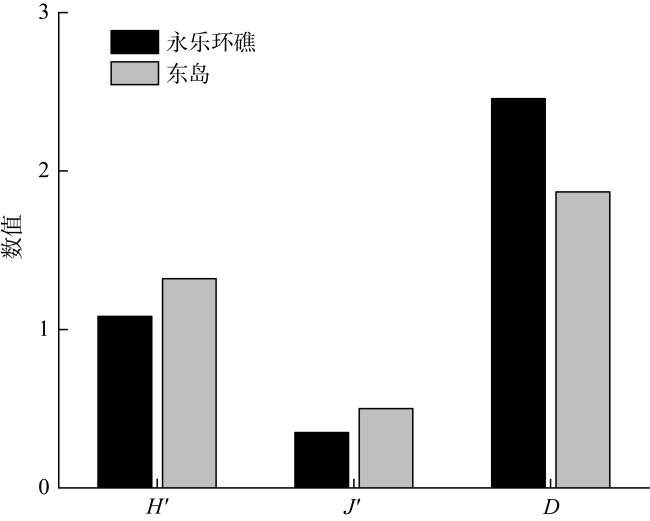
Coral reef, known as the tropical rainforest in the ocean, was an important spawning ground, nursery and feeding ground for marine fish. Yongle atoll and Dongdao Island were typical coral reef ecosystems of the Xisha Islands. In order to better manage and protect the fishery and germplasm resources of the Xisha Islands, this study collected fish eggs from the Yongle atoll and the Dongdao Island in April 2021. High-throughput sequencing technology is used to classify and identify them, and the differences in species composition and community structure between the two regions were analyzed. The results showed that only fish eggs were collected in both areas, and a total of 27 species were identified, belonging to 4 orders, 16 families and 25 genera. The perciformes was dominant, accounting for 95.45% in the Yongle atoll and 85.71% in the Dongdao Island. The similarity index of roe species composition between the Yongle atoll and the Dongdao Island was significantly different, among which the similarity of order, family, genus and species was 25.00%, 37.50%, 32.00% and 33.33%, respectively. The occurrence frequency of fish eggs was 100%. The average density of fish eggs was 0.984 ind·m-3 in the Yongle atoll and 0.453 ind·m-3 in the Dongdao Island. The dominant species (Y≥0.02) were Decapterus sp., Katsuwonus pelamis and Halichoeres trimaculatus in Yongle atoll and Scarus forsteni, Chlorurus sordidus and Parupeneus multifasciatus in the Dongdao Island, with significant differences in the dominant species. In addition, there was an undocumented species (Ambassis sp.) found in this investigation. The species richness index (D) showed that the Yongle atoll (2.46) was higher than the Dongdao Island (1.87), while Shannon-Wiener diversity index (H′) and evenness index (J′) showed that the Yongle atoll (1.08, 0.35) was lower than the Dongdao Island (1.32, 0.50). The composition of fish eggs in both regions was dominated by reef-associated fish, and the relative abundance of pelagic-oceanic fish in the Yongle atoll (88.02%) was higher than that in the Dongdao Island (0.44%). The research results showed that both the Yongle atoll and the Dongdao Island are spawning grounds for marine fish, and the Yongle atoll has an open lagoon, which is more suitable as spawning grounds for pelagic-oceanic fish (especially tuna) than the Dongdao Island. Moreover, high-throughput sequencing technology can fill the gap of traditional morphological methods in the identification of fish eggs, and could be used as a supplementary means for the investigation of fishery resources. This study provides a scientific basis for the conservation and management of fishery and germplasm resources in the Yongle atoll and the Dongdao Island.
Key words: coral reef; lagoon; Xisha Islands; spawning ground; tunas
ZHAO Jinfa , LIU Yong , LI Chunhou , WANG Teng , SHI Juan , XIAO Yayuan , WU Peng , SONG Xiaoyu . Study on species composition and distribution of fish eggs in Yongle Atoll and Dongdao Island by high-throughput sequencing technology[J]. Journal of Tropical Oceanography, 2023 , 42(6) : 127 -136 . DOI: 10.11978/2022264
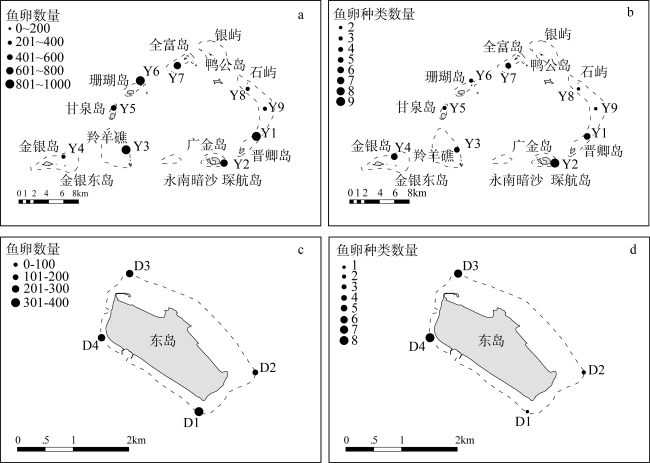
图2 永乐环礁和东岛鱼卵物种和生物量分布a. 永乐环礁鱼卵生物量; b. 永乐环礁鱼卵种类; c. 东岛鱼卵生物量; d. 东岛鱼卵种类 Fig. 2 Distribution of fish egg species and biomass in the Yongle atoll and Dongdao Island. (a) Biomass of eggs in the Yongle atoll; (b) biomass of species in the Yongle atoll; (c) number of eggs in the Dongdao Island; (d) number of species in the Dongdao Island |
表1 永乐环礁与东岛鱼卵种类组成Tab. 1 Species composition of fish eggs and larvae in the Yongle atoll and Dongdao Island |
| 目 | 科 | 属 | 种 | 生态类型 | 永乐环礁 | 东岛 | ||
|---|---|---|---|---|---|---|---|---|
| 相对丰度/% | Y | 相对丰度/% | Y | |||||
| 刺鱼目 Gasterosteiformes | 管口鱼科 Aulostomidae | 管口鱼属 Aulostomus | 中华管口鱼 A. chinensis | A | 0.00 | 0.00000 | 0.13 | 0.00031 |
| 鲈形目 Perciformes | 刺尾鱼科 Acanthuridae | 鼻鱼属 Naso | 六棘鼻鱼 N. hexacanthus | A | 0.97 | 0.00215 | 0.00 | 0.00000 |
| 突角鼻鱼 N. annulatus | A | 0.31 | 0.00068 | 0.28 | 0.00071 | |||
| 高鳍刺尾鱼属 Zebrasoma | 黄高鳍刺尾鱼 Z. flavescens | A | 0.04 | 0.00005 | 0.00 | 0.00000 | ||
| 带鱼科 Trichiuridae | 带鱼属 Trichiurus | 带鱼属未定种 Trichiurus sp. | C | 0.00 | 0.00000 | 1.02 | 0.00256 | |
| 笛鲷科 Lutjanidae | 笛鲷属 Lutjanus | 四带笛鲷 L. kasmira | A | 0.38 | 0.00043 | 0.90 | 0.00224 | |
| 蝴蝶鱼科 Chaetodontidae | 蝴蝶鱼属 Chaetodon | 叉纹蝴蝶鱼 C. auripes | A | 0.07 | 0.00015 | 0.00 | 0.00000 | |
| 隆头鱼科 Labridae | 唇鱼属 Cheilinus | 横带唇鱼 C. fasciatus | A | 0.02 | 0.00002 | 0.00 | 0.00000 | |
| 海猪鱼属 Halichoeres | 三斑海猪鱼 H. trimaculatus | A | 6.66 | 0.03700 | 0.08 | 0.00039 | ||
| 裸颊鲷科 Lethrinidae | 裸颊鲷属 Lethrinus | 桔带裸颊鲷 L. obsoletus | A | 0.02 | 0.00003 | 0.00 | 0.00000 | |
| 梅鲷科 Caesionidae | 鳞鳍梅鲷属 Pterocaesio | 黑带鳞鳍梅鲷 P. tile | A | 0.00 | 0.00000 | 6.32 | 0.01581 | |
| 鮨科 Serranidae | 侧牙鲈属 Variola | 侧牙鲈 V. louti | A | 0.06 | 0.00007 | 0.00 | 0.00000 | |
| 鲭科Scombridae | 舵鲣属 Auxis | 扁舵鲣 A. thazard | B | 0.01 | 0.00001 | 0.00 | 0.00000 | |
| 鲣属 Katsuwonus | 鲣 K. pelamis | B | 22.35 | 0.09931 | 0.00 | 0.00000 | ||
| 金枪鱼属 Thunnus | 金枪鱼属未定种 Thunnus sp. | B | 0.18 | 0.00020 | 0.00 | 0.00000 | ||
| 鲔属 Euthynnus | 鲔 E. affinis | B | 0.04 | 0.00005 | 0.00 | 0.00000 | ||
| 鲹科 Carangidae | 鲹属 Caranx | 珍鲹 C. ignobilis | A | 0.54 | 0.00120 | 0.00 | 0.00000 | |
| 圆鲹属 Decapterus | 圆鲹属未定种 Decapterus sp. | B | 65.44 | 0.50900 | 0.35 | 0.00087 | ||
| 双边鱼科 Ambassidae | 双边鱼属 Ambassis | 双边鱼属未定种 Ambassis sp. | B | 0.00 | 0.00000 | 0.09 | 0.00024 | |
| 羊鱼科 Mullidae | 副绯鲤属 Parupeneus | 多带副绯鲤 P. multifasciatus | A | 0.03 | 0.00003 | 8.49 | 0.04247 | |
| 拟羊鱼属 Mulloidichthys | 无斑拟羊鱼 M. vanicolensis | A | 0.30 | 0.00034 | 0.00 | 0.00000 | ||
| 鹦嘴鱼科 Scaridae | 大鹦嘴鱼属 Bolbometopon | 隆头大鹦嘴鱼 B. muricatum | A | 0.37 | 0.00041 | 0.06 | 0.00016 | |
| 绿鹦嘴鱼属 Chlorurus | 灰鹦嘴鱼 C. sordidus | A | 0.38 | 0.00085 | 16.94 | 0.08470 | ||
| 鹦嘴鱼属 Scarus | 绿唇鹦嘴鱼 S. forsteni | A | 1.35 | 0.00300 | 60.24 | 0.30120 | ||
| 许氏鹦嘴鱼 S. schlegeli | A | 0.01 | 0.00001 | 3.92 | 0.00979 | |||
| 鼠鱚目 Gonorhynchiformes | 遮目鱼科 Chanidae | 遮目鱼属 Chanos | 遮目鱼 C. chanos | C | 0.45 | 0.00050 | 0.00 | 0.00000 |
| 鲀形目 Tetraodontiformes | 单角鲀科 Monacanthidae | 前角鲀属 Pervagor | 前角鲀属未定种 Pervagor sp. | A | 0.00 | 0.00000 | 1.18 | 0.00295 |
注: A. 礁栖鱼类; B. 大洋性中上层鱼类; C. 深海鱼类 |
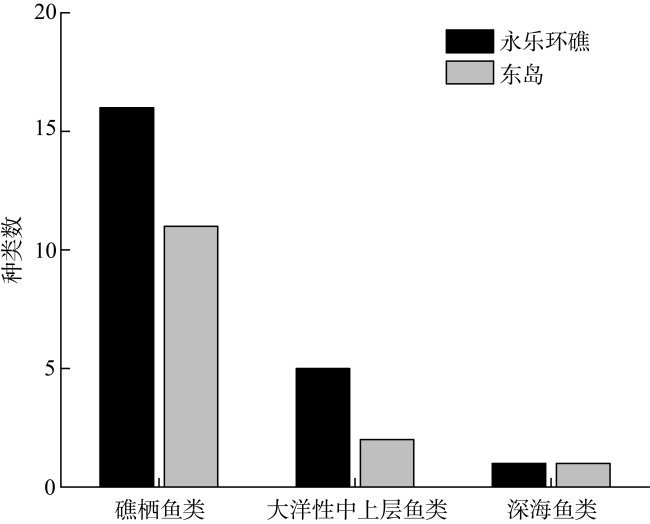
图3 永乐环礁和东岛鱼卵不同生态类型的种类组成Fig. 3 Species composition of different ecological types of fish eggs in the Yongle atoll and Dongdao Island |
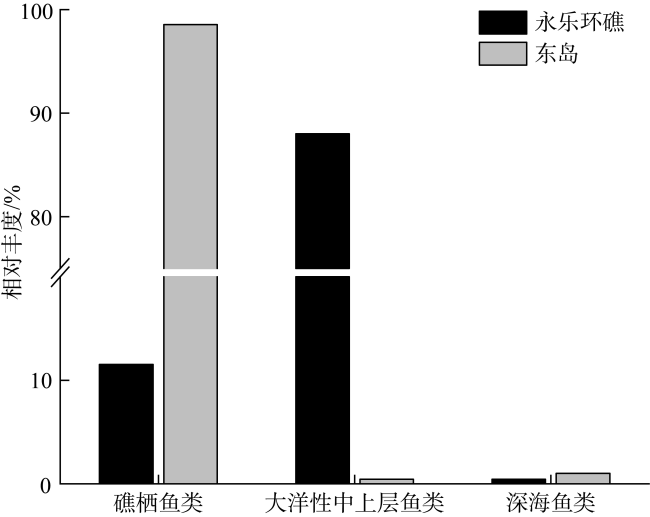
图4 永乐环礁和东岛鱼卵不同生态类型的相对丰度Fig. 4 Relative abundance of different ecological types of fish eggs in the Yongle atoll and Dongdao Island |
| [1] |
陈真然, 1979. 西沙、中沙群岛海域的浮性鱼卵和仔稚鱼[J]. 水产科技情报, (4): 11-13.
|
| [2] |
陈真然, 魏淑珍, 1978. 西沙、中沙群岛海域浮性鱼卵和仔稚鱼的初步调查研究[R]// 中国科学院南海海洋研究所. 我国西沙、中沙群岛海域海洋生物调查研究报告集. 北京: 科学出版社, 295-320.
|
| [3] |
郭琴, 高雷, 潘文杰, 等, 2020. 赣江下游丰城段鱼类早期资源现状调查[J]. 水生态学杂志, 41(6): 106-112.
|
| [4] |
胡兴坤, 邵科, 阙延福, 等, 2022. 金沙江下游宜宾段产漂流性卵鱼类早期资源动态变化[J]. 水生态学杂志, 44(2): 73-80.
|
| [5] |
李晓敏, 马毅, 张杰, 等, 2020. 基于高分遥感的西沙群岛典型海岛时空变化分析[J]. 海洋通报, 39(6): 717-729.
|
| [6] |
李永振, 贾晓平, 陈国宝, 2007. 南海珊瑚礁鱼类资源[M]. 北京: 海洋出版社.
|
| [7] |
李永振, 史赟荣, 艾红, 等, 2011. 南海珊瑚礁海域鱼类分类多样性大尺度分布格局[J]. 中国水产科学, 18(3): 619-628.
|
| [8] |
卢芷程, 李敏, 张俊, 等, 2021. 基于DNA条形码的南海美济礁潟湖鱼卵种类鉴定与组成初探[J]. 南方水产科学, 17(6): 12-21.
|
| [9] |
马红英, 张晗, 邓捷, 等, 2021. 山溪鲵属的水样环境DNA分析方法的建立[J]. 安徽农业大学学报, 48(5): 784-789.
|
| [10] |
邱书婷, 刘昕明, 陈彬, 等, 2022. 西沙群岛珊瑚礁鱼类多样性及分布格局[J]. 海洋环境科学, 41(3): 395-401.
|
| [11] |
舒璐, 林佳艳, 徐源, 等, 2020. 基于环境DNA宏条形码的洱海鱼类多样性研究[J]. 水生生物学报, 44(5): 1080-1086.
|
| [12] |
孙典荣, 林昭进, 邱永松, 等, 2005. 西沙群岛重要珊瑚礁海域鱼类区系[J]. 南方水产科学, 1(5): 18-25.
|
| [13] |
万瑞景, 张仁斋, 2016. 中国近海及其邻近海域鱼卵和仔稚鱼[M]. 上海: 上海科学技术出版社.
|
| [14] |
王腾, 刘永, 李纯厚, 等, 2022a. 永兴岛附近海域珊瑚礁鱼类群落结构特征[J]. 水生生物学报, 47(4): 674-683.
|
| [15] |
王腾, 刘永, 全秋梅, 等, 2022b. 七连屿珊瑚礁鱼类种类组成特征分析[J]. 中国水产科学, 29(1): 102-117.
|
| [16] |
吴娜, 张楠, 曹明, 等, 2018. 基于DNA条形码技术的永暑礁泻湖鱼卵鉴定研究[J]. 淡水渔业, 48(2): 51-57.
|
| [17] |
肖瑜璋, 王蓉, 郑琰晶, 等, 2013. 珠江口鱼类浮游生物种类组成与数量分布[J]. 热带海洋学报, 32(6): 80-87.
|
| [18] |
徐华, 王斌, 张聪, 等, 2021. 黄河口近岸海域鱼卵、仔稚鱼种类组成及群落结构特征[J]. 海洋科学, 45(11): 105-117.
|
| [19] |
闫欣, 钟俊生, 王明星, 等, 2014. 杭州湾北部水域仔稚鱼种类组成和多样性研究[J]. 上海海洋大学学报, 23(5): 765-773.
|
| [20] |
闫亚利, 2019. 南海部分岛礁鱼卵及成鱼的DNA条形码研究[D]. 上海: 上海海洋大学.
|
| [21] |
张俊彬, 黄增岳, 2003. 阳江东平核电站邻近海区鱼卵和仔鱼调查研究[J]. 热带海洋学报, 22(3): 78-84.
|
| [22] |
张仁斋, 1983. 三种金枪鱼类(鲣、黄鳍金枪鱼、扁舵鲣)的仔、稚鱼在南海的分布和产卵期[J]. 海洋学报(中文版), 5(3): 368-375.
|
| [23] |
张婷, 林柳, 蹇丽, 等, 2020. 西沙群岛七连屿绿海龟(Chelonia mydas)产卵场海滩垃圾调查[J]. 生态学杂志, 39(7): 2408-2415.
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}