
Journal of Tropical Oceanography >
Effects of seawater acidification on microorganisms associated with Aurelia coerulea polyps
Copy editor: LIN Qiang
Received date: 2023-02-01
Revised date: 2023-03-28
Online published: 2023-04-07
Supported by
National Natural Science Foundation of China(41876138)
National Natural Science Foundation of China(U2106208)
Strategic Priority Research Program of the Chinese Academy of Sciences(XDA23050301)
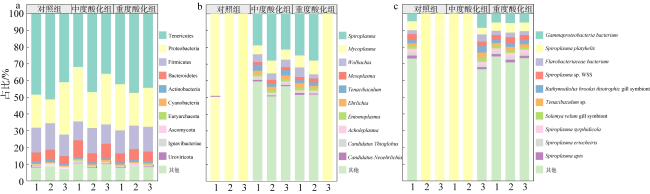
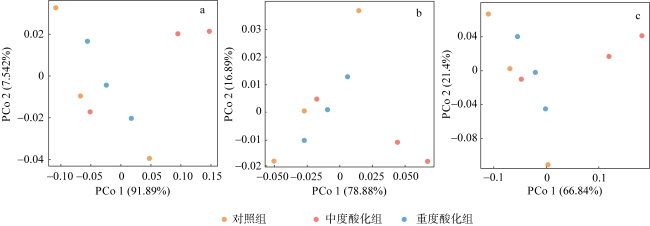
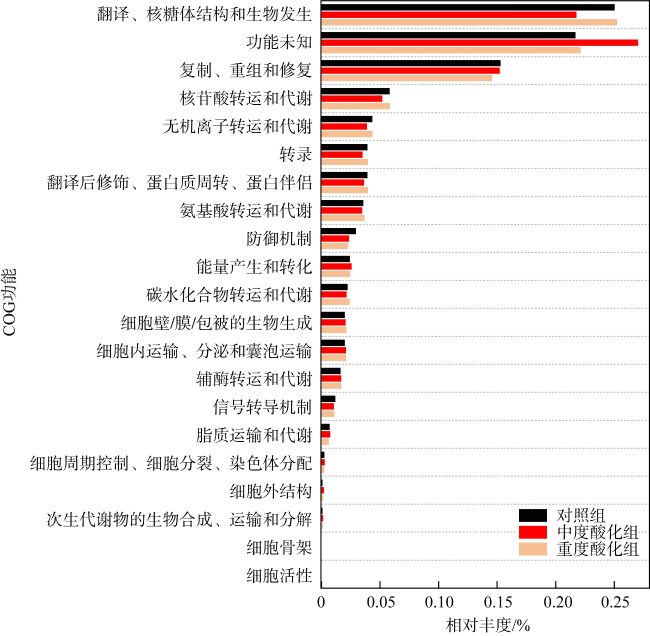
Ocean acidification (OA) poses strong threat to marine organisms, and studying the impact of OA on their associated microbial communities is important for understanding the adaptive mechanisms of host and microbial response to environmental stresses. Moon jellyfish (Aurelia coerulea) is the dominant species of disaster jellyfish in China and plays an important role in marine ecosystem. In this study, we investigated the effects of future OA (pH 7.8 and 7.6) on the structure and function of the A. coerulea polyp-associated microbial communities using Biolog-ECO and metagenomic approaches. The results showed that the structures and functions of the microbial community associated with A. coerulea polyps were conserved in different degrees of seawater acidification. Biolog-ECO results showed that seawater acidification had little effect on the microbial metabolic activity, diversity index and carbon source utilization of the associated microbial community of A. coerulea polyps. The metagenomic results showed that seawater acidification did not affect the dominant species and community structure of the microorganisms. Tenericute, Proteobacteria, Firmicutes and Bacteroidetes were the dominant groups of the community. Moreover, there was no significant difference in the function of associated microorganisms exposed to different acidification conditions. Therefore, we speculate that future OA will not affect the polyps by altering the community structures of their associated microorganisms.
SUN Tingting , HAO Wenjin , XU Pengzhen , YE Lijing , DONG Zhijun . Effects of seawater acidification on microorganisms associated with Aurelia coerulea polyps[J]. Journal of Tropical Oceanography, 2023 , 42(6) : 111 -119 . DOI: 10.11978/2023011
表1 暴露实验测量计算海水碳酸盐化学参数Tab. 1 Measured and calculated carbonate chemistry parameters of seawater for the exposure experiment |
| 处理组 | pH | 温度/℃ | 盐度/‰ | TA/(μmol·kg-1) | pCO2/μatm | DIC/(μmol·kg-1) |
|---|---|---|---|---|---|---|
| 对照 | 8.14±0.02 | 21.0±0.0 | 28.6±0.9 | 2521.53±34.21 | 529.67±27.56 | 2307.44±29.02 |
| 中度酸化 | 7.80±0.00 | 21.0±0.0 | 28.6±0.8 | 2523.60±36.20 | 1252.19±15.65 | 2447.38±33.77 |
| 重度酸化 | 7.60±0.01 | 21.0±0.0 | 28.5±0.8 | 2494.88±31.88 | 2028.32±28.00 | 2485.80±30.12 |
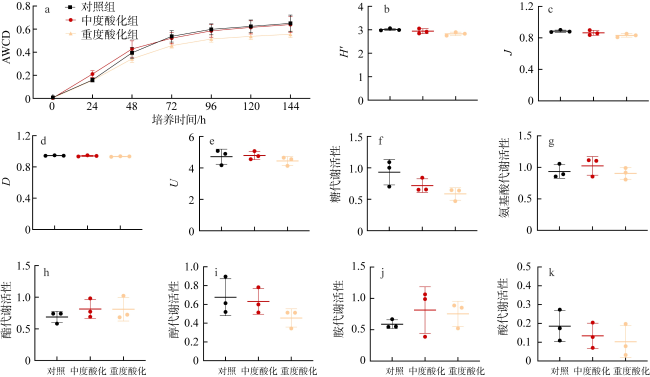
图1 海月水母螅状体共附生微生物代谢活性a. AWCD变化曲线; b—e. 微生物多样性指数; f—k. 微生物对6类碳源的利用 Fig. 1 Metabolic activity of symbiotic and epibiotic microorganism in A. coerulea polyps. (a) AWCD change curve; (b-e) microbial diversity index; (f-k) microbial utilization of class 6 carbon sources |
表2 基于 Biolog-ECO 技术的海月水母螅状体共附生微生物AWCD、多样性指数和6类碳源利用的单因素方差分析结果Tab. 2 One-way ANOVA results of AWCD, diversity index and utilization of 6-type carbon sources by microorganisms associated with A. coerulea polyps based on Biolog-ECO technique |
| 测量指标 | F | P | |
|---|---|---|---|
| AWCD | 144h | F2,6=2.287 | P=0.183 |
| 多样性指数 | Shannon's指数(H') | F 2,6=4.008 | P=0.078 |
| Pielou指数(J) | F 2,6=4.008 | P=0.078 | |
| Simpson指数(D) | F 2,6=4.426 | P=0.066 | |
| Mclntosh指数(U) | F 2,6=0.794 | P=0.495 | |
| 碳源 | 糖 | F 2,6=4.356 | P=0.068 |
| 氨基酸 | F 2,6=0.839 | P=0.477 | |
| 酯 | F 2,6=0.713 | P=0.528 | |
| 醇 | F 2,6=1.828 | P=0.240 | |
| 胺 | F 2,6=0.670 | P=0.546 | |
| 酸 | F 2,6=0.837 | P=0.478 | |
| [1] |
高尚武, 洪惠馨, 张士美, 2002. 中国动物志无脊椎动物第27卷[M]. 北京: 科学出版社 (in Chinese).
|
| [2] |
马克平, 黄建辉, 于顺利, 等, 1995. 北京东灵山地区植物群落多样性的研究: Ⅱ丰富度、均匀度和物种多样性指数[J]. 生态学报, 15(3): 268-277.
|
| [3] |
汤毓祥, 孙洪亮, 胡筱敏, 等, 2007. 海洋调查规范第2部分: 海洋水文观测: GB/T 12763.2-2007[S]. 北京: 中华人民共和国国际质量监督检验检疫总局, 中国国家标准化管理委员会: 3-7.
|
| [4] |
袁翔城, 梁宇娴, 宋严, 等, 2023. CO2升高对风信子鹿角珊瑚(Acropora hyacinthus)钙化速率和基因表达的影响[J/OL]. 热带海洋学报. [2023-02-01]. http://www.jto.ac.cn/CN/10.11978/2022182.
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
MILLERO,
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}