
Journal of Tropical Oceanography >
Difference in the feeding contents of three hermit crabs in the coral reefs of the Nansha Islands, South China Sea
Copy editor: LIN Qiang
Received date: 2023-03-07
Revised date: 2023-04-17
Online published: 2023-04-22
Supported by
National Natural Science Foundation of China(42176118)
National Key Research and Development Project of China(2021YFC3100500)
Science and Technology Planning Project of Guangdong Province, China(2020B1212060058)
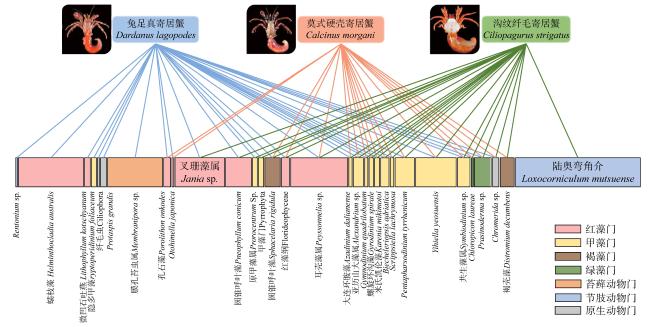
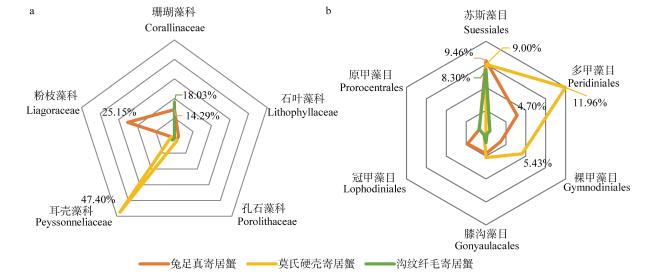
Hermit crab is a type of marine benthic animal. The complex structure of coral reefs makes it one of the areas with the highest diversity of hermit crabs. In response to changes in environmental conditions, such as increasing coverage of benthic algae, small benthic organisms such as hermit crabs can quickly adapt to changes in food resources and play a similar function to herbivorous fishes in maintaining the health of coral reef ecosystems. However, little is known about the differences in food composition on site and ecological functional positioning of different hermit crab species. In this study, three dominant hermit crab species were collected from the coral reef of the Nansha Islands in the spring of 2019: Dardanus lagopodes, Calcinus morgani, and Ciliopagurus strigatus, and their on-site food composition was analyzed using high-throughput sequencing technology. The results showed that there were nine phyla of food types for the three species of hermit crabs, including Rhodophyta, Pyrrophyta, Phaeophyta, Chlorophyta, Arthropoda and Bryozoa, etc. All three hermit crab species fed mainly on algae, including large amounts of Rhodophyta and Pyrrophyta algae (37.75% and 21.14%), and small amounts of Chlorophyta algae (3.13%). But the food composition of different hermit crab species was specific. Calcinus morgani fed mainly on algae turf and detritus and was a herbivorous hermit crab. Ciliopagurus strigatus took Ostracoid from epilithic algal matrix (EAM) as the main food source, accompanied by some branching coralline algae and filamentous Phaeophyta algae. Dardanus lagopodes fed on crustose coralline algae (CCA), macroalgae and epiphytic bryozoans. The results revealed obvious differences in the main foods of marine hermit crabs. Combined with the analysis of ecological niche overlap, it is found that the overlap of the three species of hermit crabs was low (0.059). The feeding areas of Dardanus lagopodes and the other two hermit crab species may not overlap in degraded coral reef areas dominated by CCA. Dardanus lagopodes plays an important role in clearing CCA and bryozoans, and maintaining the healthy development of coral reefs. Ciliopagurus strigatus and Calcinus morgani, as secondary consumers, transfer energy more efficiently to the upper level. These research results extend the previous understanding of the ecological status of hermit crabs.
Key words: hermit crab; diet; high-throughput sequencing; ecological function; coral reef
JIA Nan , ZHOU Tiancheng , HU Simin , ZHANG Chen , HUANG Hui , LIU Sheng . Difference in the feeding contents of three hermit crabs in the coral reefs of the Nansha Islands, South China Sea[J]. Journal of Tropical Oceanography, 2024 , 43(3) : 109 -121 . DOI: 10.11978/2023029
表1 寄居蟹样品的相关信息Tab. 1 Information about hermit crab samples |
| 种名 | 盾长/mm | 螺壳宽/mm | 螺长/mm | 螺口宽/mm | 寄居蟹湿重/g | 个体数 |
|---|---|---|---|---|---|---|
| 兔足真寄居蟹Dardanus lagopodes | 8.22±2.23 | 62.74±39.28 | 48.55±18.31 | 12.18±7.50 | 3.98±2.76 | 5 |
| 莫氏硬壳寄居蟹Calcinus morgani | 4.79±1.23 | 23.29±4.60 | 33.42±7.30 | 4.48± 1.52 | 0.58±0.47 | 5 |
| 沟纹纤毛寄居蟹Ciliopagurus strigatus | 4.32 | 18.82 | 31.54 | 1.90 | 0.38 | 2 |
图3 基于三种寄居蟹肠道内容物18S rDNA序列构建的系统进化树Fig. 3 Maximum likelihood tree of 18S rDNA sequence from hermit crab gut contents |
表2 三种寄居蟹的营养生态位宽度Tab. 2 Trophic niches breadth of the three species of hermit crabs |
| 种名 | 营养生态位宽度 |
|---|---|
| 兔足真寄居蟹 Dardanus lagopodes | 7.67 |
| 莫氏硬壳寄居蟹 Calcinus morgani | 4.26 |
| 沟纹纤毛寄居蟹 Ciliopagurus strigatus | 2.62 |
图6 三种寄居蟹食物来源的食物网物种图片引用自McLaughlin等(2007) Fig. 6 Networks depicting the main dietary complementary and overlaps of the three species of hermit crabs |
表3 寄居蟹间的生态位重叠度Tab. 3 Niches overlap degree between hermit crabs |
| 寄居蟹种类 | 兔足真 寄居蟹 | 莫氏硬壳 寄居蟹 | 沟纹纤毛 寄居蟹 |
|---|---|---|---|
| 兔足真寄居蟹 | — | 0.071 | 0.043 |
| 莫氏硬壳寄居蟹 | 0.071 | — | 0.062 |
| 沟纹纤毛寄居蟹 | 0.043 | 0.062 | — |
| [1] |
蔡慧梅, 1982. 南海东北部海区介形虫的分布[J]. 热带海洋, 1(1): 42-57.
|
| [2] |
韩源源, 2017. 中国海陆生寄居蟹科和寄居蟹科(甲壳动物亚门: 异尾下目)的系统分类学研究[D]. 太原: 山西师范大学:163-167.
|
| [3] |
黄海燕, 陆斗定, 夏平, 等, 2009. 2006年冬季长江口海域表层沉积物中甲藻孢囊的分类学研究[J]. 生态学报, 29(11): 5902-5911.
|
| [4] |
康伟, 王朝晖, 2016. 桂山岛海域甲藻孢囊的分布与浮游植物休眠体萌发研究[J]. 海洋科学, 40(7): 33-40.
|
| [5] |
李月, 王云龙, 欧阳珑玲, 2022. 浮游虫黄藻生长及碱性磷酸酶对不同磷酸盐浓度的响应[J]. 海洋渔业, 44(4): 459-467.
|
| [6] |
廖彤晨, 尹健强, 李开枝, 等, 2019. 海洋浮游介形类的多样性和生态学研究进展[J]. 生态科学, 38(6): 190-198.
|
| [7] |
廖芝衡, 2021. 南海珊瑚群落和底栖海藻的空间分布特征及其生态影响[D]. 南宁: 广西大学:84-86.
|
| [8] |
林先智, 胡思敏, 刘胜, 等, 2018. 传统测序与高通量测序在稚鱼食性分析中的比较[J]. 应用生态学报, 29(9): 3093-3101.
|
| [9] |
刘会莲, 刘锡兴, 2016. 中国海洋苔藓动物分布特点与区系性质初探[M]// 国际生物多样性计划中国委员会, 环境保护部, 中科院, 中国生物多样性保护与研究进展Ⅸ. 北京: 气象出版社: 61-72 (in Chinese).
|
| [10] |
刘宇洋, 2020. 三种有害藻华甲藻产休眠孢囊的证据及其在中国近海沉积物中的分布[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 69-82.
|
| [11] |
潘子良, 2017. 黄岩岛造礁石珊瑚共生藻密度的种间、空间差异及其生态意义[D]. 南宁: 广西大学: 1-3.
|
| [12] |
王复振, 1992. 中国寄居蟹类区系研究[J]. 东海海洋, 10(1): 59-63.
|
| [13] |
王翔宇, 詹冬梅, 李美真, 等, 2011. 大型海藻吸收氮磷营养盐能力的初步研究[J]. 渔业科学进展, 32(4): 67-71.
|
| [14] |
王朝晖, 张宇宁, 王文婷, 等, 2022. 福建东山湾表层沉积物中甲藻孢囊分布研究[J]. 热带海洋学报, 41(4): 154-162.
|
| [15] |
肖丽婵, 2013. 中国海活额寄居蟹科(Diogenidae)系统分类学研究[D]青岛: 中国科学院研究生院(海洋研究所): 211-221.
|
| [16] |
徐林通, 郑艳坤, 郝俊, 等, 2018. 珊瑚共生虫黄藻研究进展[J]. 河北渔业, (8): 56-59, 62.
|
| [17] |
张琛, 胡思敏, 林先智, 等, 2022. 南沙珊瑚礁区波纹钩鳞鲀(Balistapus undulatus)食性和营养级分析[J]. 热带海洋学报, 41(1): 7-14.
|
| [18] |
张德瑞, 周锦华, 1981. 耳壳藻科一新属——枝壳藻属[J]. 海洋与湖沼, 12(6): 538-544.
|
| [19] |
张松龄, 刘锡兴, 1995. 中国南部水域膜孔苔虫属一新种(苔藓动物门: 唇口目: 膜孔苔虫科)[J]. 动物分类学报, 20(2): 133-136.
|
| [20] |
周天成, 2020. 三亚鹿回头近岸珊瑚礁区寄居蟹的摄食及生态功能探讨[D]. 北京: 中国科学院大学: 1-18.
|
| [21] |
周天成, 胡思敏, 林先智, 等, 2020. 基于18S rDNA条形码技术的珊瑚礁区塔形马蹄螺(Tectus pyramis)食性分析[J]. 海洋科学, 44(2): 99-107.
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}