
Journal of Tropical Oceanography >
Development of gonads with annual cycle in Anlactinia sinensis
Copy editor: LIN Qiang
Received date: 2023-02-17
Revised date: 2023-04-17
Online published: 2023-04-23
Supported by
Ocean University of China-Student Research Development Program(202110423574X)
The study on animal gonad development can reveal the process of gametogenesis, and evaluate the fertility and source utilization of a species. The Chinese tube anemone Aulactinia sinensis is a dominant species in intertidal zone of the Yellow Sea, China. In this study, we reported characteristics of the gonad development with annual cycle in A. sinensis inhabiting Huiquan Bay, Qingdao. The results showed that A. sinensis is cylindrical in appearance, and the tentacle arrangement of oral disc is 6+6+12+24+48. There are 48 pairs of mesenteries inside the column. A. sinensis is dioecism, and populational ratio of female to male is 1.25: 1. During the reproductive season, all mesenteries can develop into reproductive mesenteries, and form the gonad. According to the characteristics of spermatogenicvesicle, germ cell types, and composition in the mesenteries, the development process of gonads was divided into four stages, and the gamete spawning occurred between May to September. The fecundity of an individual with the oral disc diameter of 3 cm was 188277.91 ± 28861.91. Our findings will provide basic scientific data for studying the source recycling of A. sinensis.
Key words: Aulactinia sinensis; morphology; gonad; annual development; fecundity
LI Yunpeng , WANG Zhicheng , ZHANG Long , ZHANG Wei , DU Mengda , SUN Yi , ZHANG Zhifeng . Development of gonads with annual cycle in Anlactinia sinensis[J]. Journal of Tropical Oceanography, 2023 , 42(6) : 120 -126 . DOI: 10.11978/2023019
图 2 中华管海葵的内部结构a. 具口道壁的消化循环腔; b. 去除口道壁的消化循环腔。1: 触手; 2: 口道沟; 3: 口道壁; 4: 初级隔膜; 5: 次级隔膜; 6: 三级隔膜; 7: 特化后生殖腺; 8: 隔膜丝; 9: 体壁; 10: 指向隔膜 Fig. 2 Internal structure of Aulactinia sinensis. (a) Gastrovascular cavity with stomodaeum wall; (b) gastrovascular cavity without stomodaeum wall. (1) tentacle; (2) stomodaeum groove; (3) stomodaeum wall; (4) primary mesentery; (5) secondary mesentery; (6) tertiary mesentery; (7) specialized reproductive mesentery; (8) mesenterial filament; (9) wall; (10) directive mesentery |
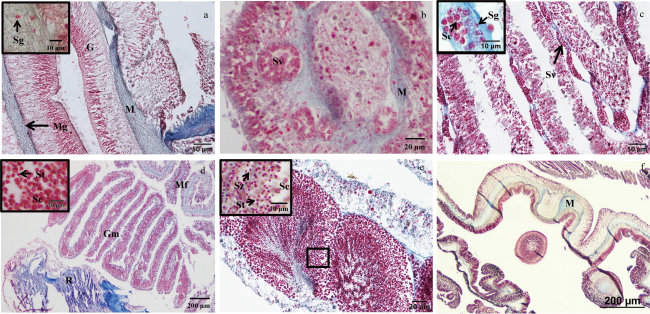
图 3 中华管海葵雄性生殖隔膜组织学结构的年周期变化a. 12月; b. 2月; c. 3月; d. 4月; e. 7月; f. 11月。G: 胃层; Gm: 生殖隔膜; M: 中胶层; Mf: 隔膜丝; Mg: 接近中胶层的生殖细胞; R: 弥散性牵引肌; Sc: 精母细胞; Sg: 精原细胞; St: 精细胞; Sv: 精小囊; Sz: 精子 Fig. 3 Histological structure of the reproductive mesenteries with annual cycle in Aulactinia sinensis males. (a) December; (b) February; (c) March; (d) April; (e) July; (f) November. G: gastrodermis; Gm: genital mesentery; M: mesoglea; Mg: germ cell close with mesoglea; Mf: mesenteric filamen; R: diffuse mesenterial retractor muscle; Sc: spermatocyte; Sg: spermatogonium; St: sperm cell; Sv: spermatogenicvesicle; Sz: sperm |
图4 中华管海葵雌性生殖隔膜组织学结构的年周期变化a. 1月; b. 2月; c. 5月; d. 7月; e.10月; f. 11月。G: 胃层; Gm: 生殖隔膜; M: 中胶层; N: 细胞核; Nu: 核仁; Oc: 卵母细胞; Ol: 卵巢小叶; Oo: 卵原细胞 Fig. 4 Histological structure of the reproductive mesenteries with annual cycle in Aulactinia sinensis females. (a) January; (b) February; (c) May; (d) July; (e) October. (f) November. G: gastrodermis; Gm: genital mesentery; M: mesoglea; N: nucleus; Nu: nucleolus; Oc: oocyte; Ol: ovarian lobule; Oo: oogonium. |
表1 中华管海葵的繁殖力Tab. 1 The fecundity of Aulactinia sinensis |
| 统计指标 | 平均值±标准差 | 相对范围 |
|---|---|---|
| 可育口盘直径/cm (n=36) | 3.81±0.95 | 2.30~6.00 |
| 卵细胞直径/μm (n=3) | 96.63±9.85 | 74.74~125.32 |
| 单条初级隔膜怀卵量/个 (n=30) | 188277.91±28861.91 | 115255.94~233263.63 |
| [1] |
|
| [2] |
陈介康, 1986. 腔肠动物标本的麻醉方法[J]. 生物学通报, (8): 42-43 (in Chinese).
|
| [3] |
李霞, 贾玉妹, 李雅娟, 2004. 黄海葵解剖学和主要器官组织学的研究[J]. 大连海洋大学学报, 19(3): 171-175.
|
| [4] |
李阳, 徐奎栋, 2020. 中国海海葵目(刺胞动物门: 珊瑚虫纲)物种多样性与区系特点[J]. 海洋与湖沼, 51(3): 434-443.
|
| [5] |
李元超, 黄晖, 董志军, 等, 2009. 鹿回头佳丽鹿角珊瑚卵母细胞发育的组织学研究[J]. 热带海洋学报, 28(1): 56-60.
|
| [6] |
申玉春, 杨小东, 刘丽, 等, 2014. 澄黄滨珊瑚、大管孔珊瑚和丛生盔形珊瑚性腺发育与繁殖力的研究[J]. 海洋与湖沼, 45(2): 314-321.
|
| [7] |
杨海萍, 王储庆, 吴建平, 等, 2015. 几种麻醉剂对中华仙影海葵麻醉效果的比较[J]. 水产养殖, 36(4): 8-13.
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}